

System-wide Benefits of Intermeal Fasting by Autophagy
- PMID: 29107505
- PMCID: PMC5718973
- DOI: 10.1016/j.cmet.2017.09.020
System-wide Benefits of Intermeal Fasting by Autophagy
Abstract
Autophagy failure is associated with metabolic insufficiency. Although caloric restriction (CR) extends healthspan, its adherence in humans is poor. We established an isocaloric twice-a-day (ITAD) feeding model wherein ITAD-fed mice consume the same food amount as ad libitum controls but at two short windows early and late in the diurnal cycle. We hypothesized that ITAD feeding will provide two intervals of intermeal fasting per circadian period and induce autophagy. We show that ITAD feeding modifies circadian autophagy and glucose/lipid metabolism that correlate with feeding-driven changes in circulating insulin. ITAD feeding decreases adiposity and, unlike CR, enhances muscle mass. ITAD feeding drives energy expenditure, lowers lipid levels, suppresses gluconeogenesis, and prevents age/obesity-associated metabolic defects. Using liver-, adipose-, myogenic-, and proopiomelanocortin neuron-specific autophagy-null mice, we mapped the contribution of tissue-specific autophagy to system-wide benefits of ITAD feeding. Our studies suggest that consuming two meals a day without CR could prevent the metabolic syndrome.
Keywords: POMC; aging; autophagy; caloric restriction; circadian; fatty liver; gluconeogenesis; metabolic syndrome; myogenic progenitors; twice-a-day feeding.
Copyright © 2017 Elsevier Inc. All rights reserved.
Figures
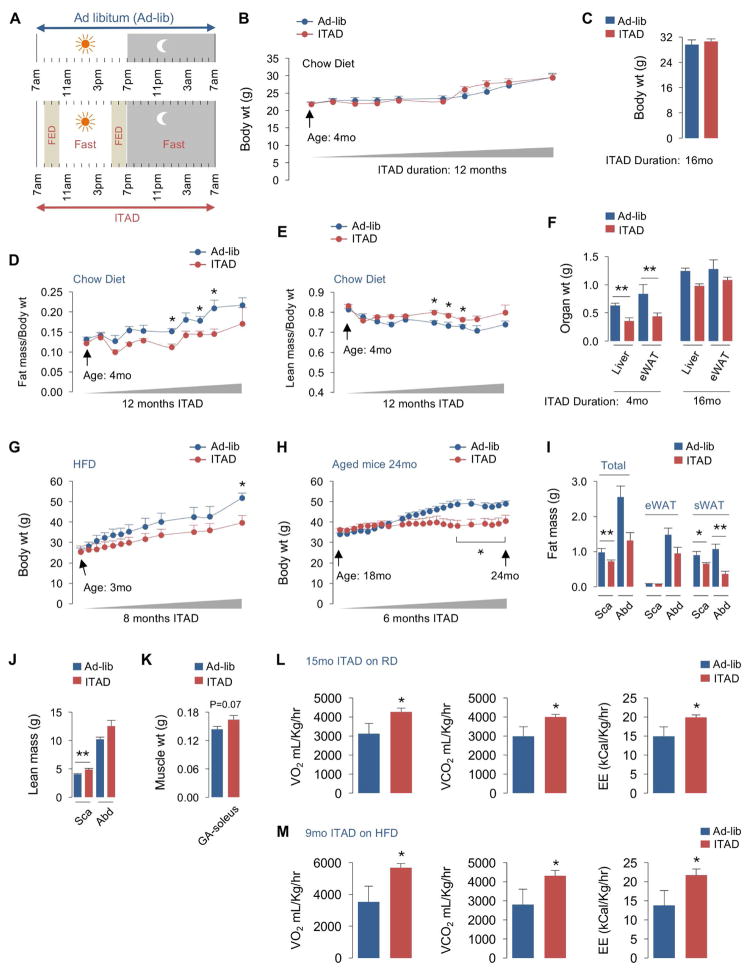
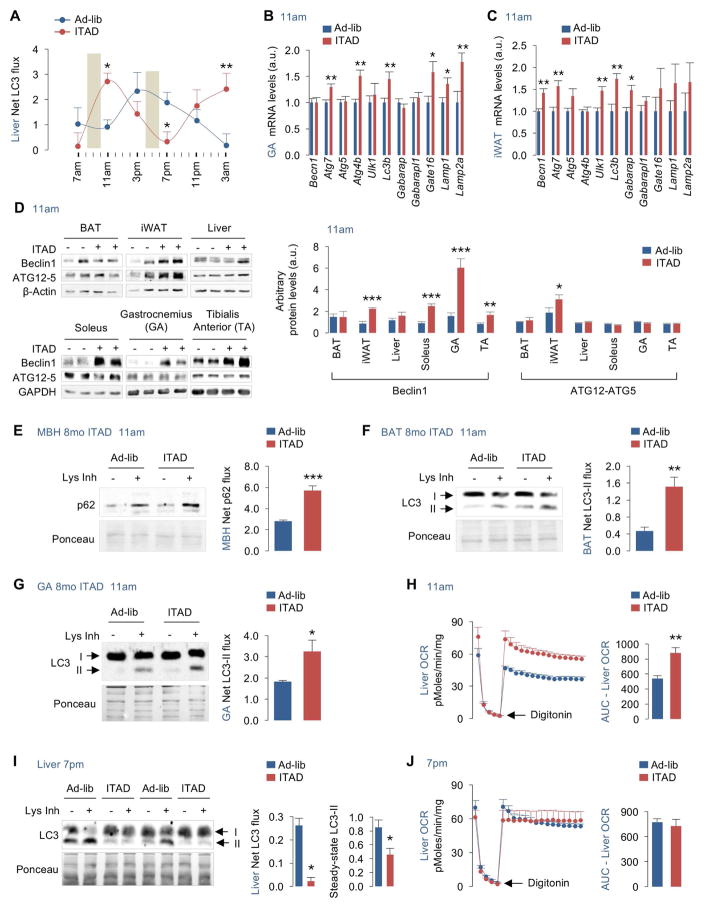
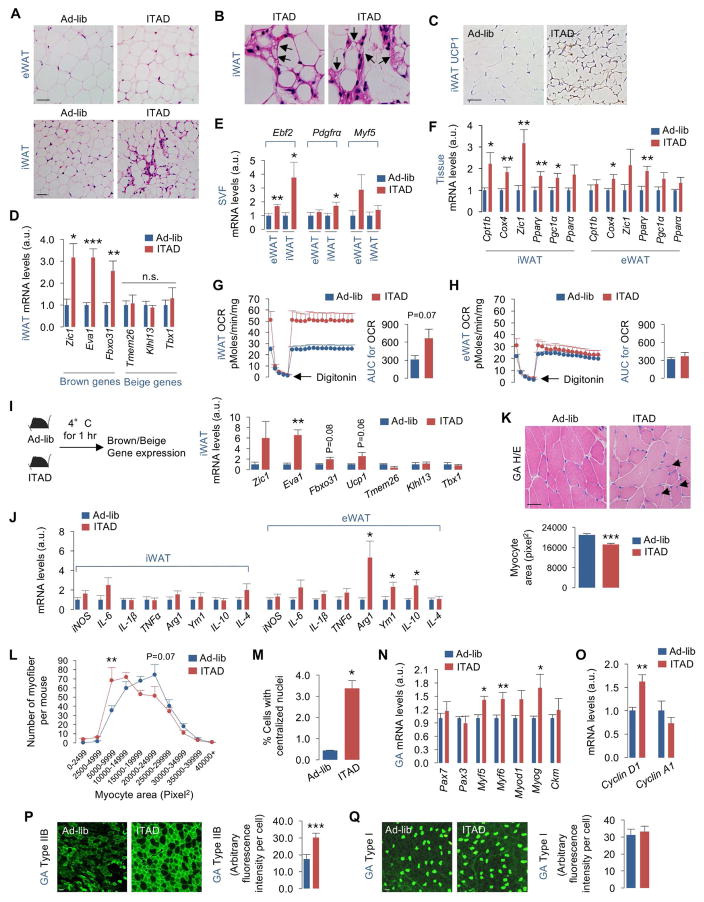
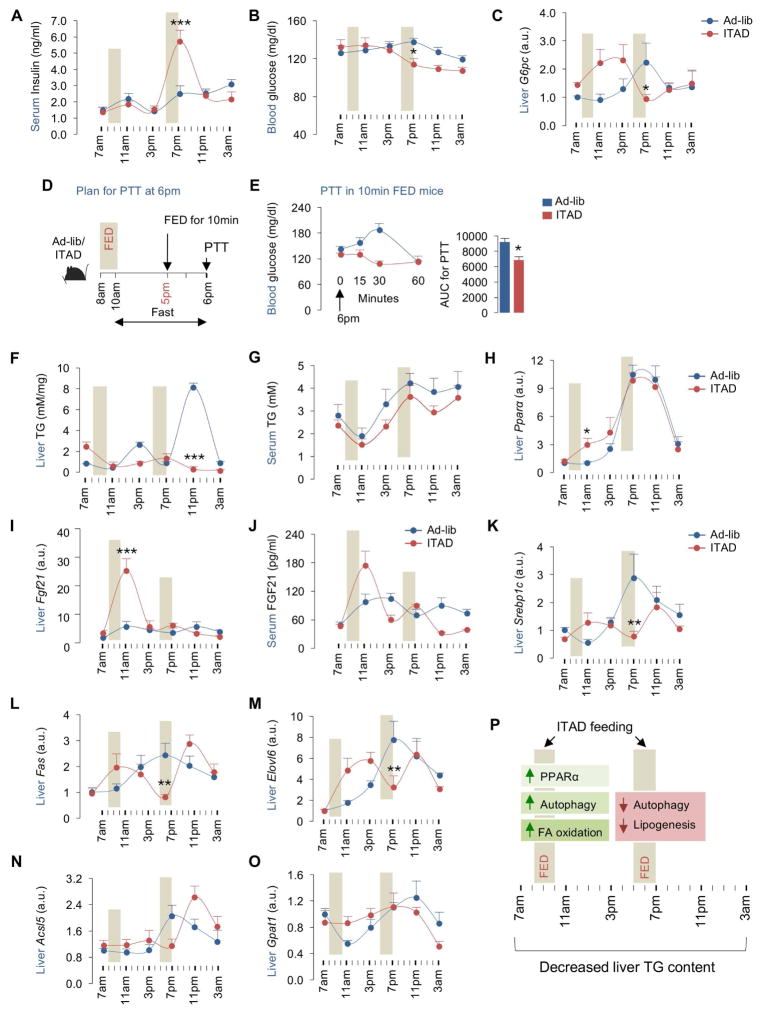
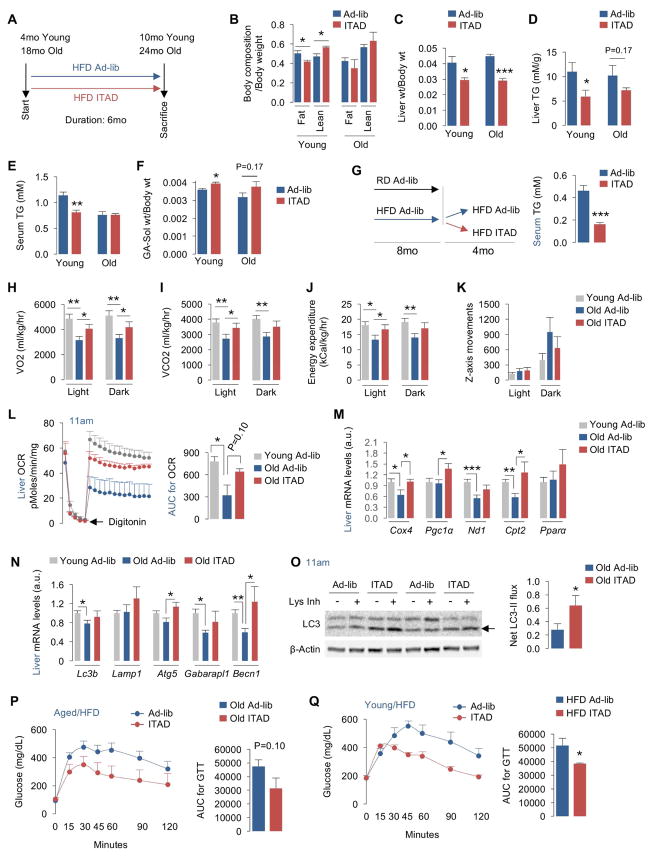
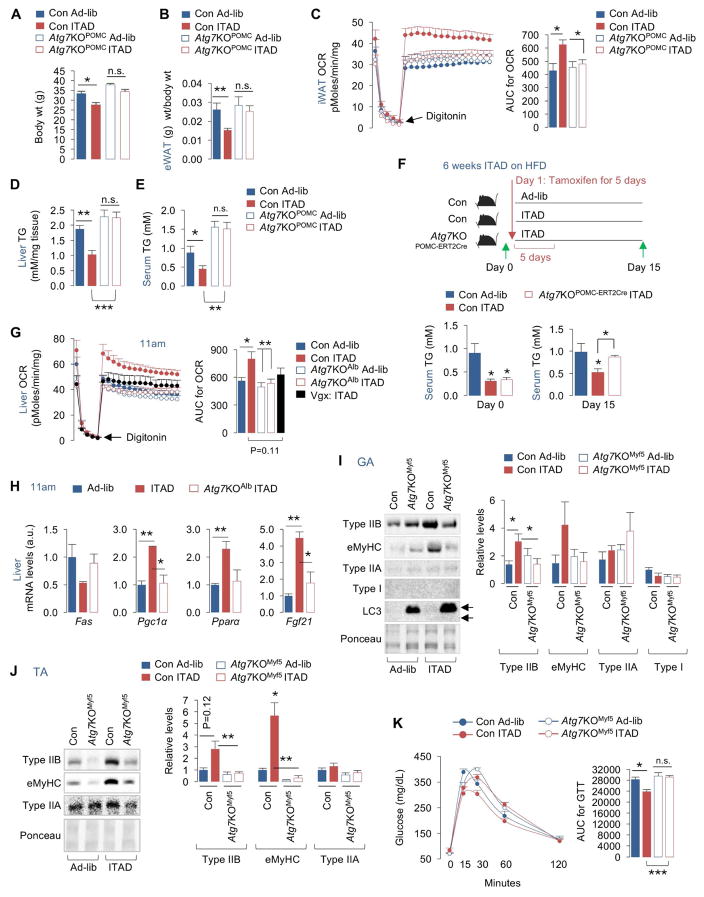
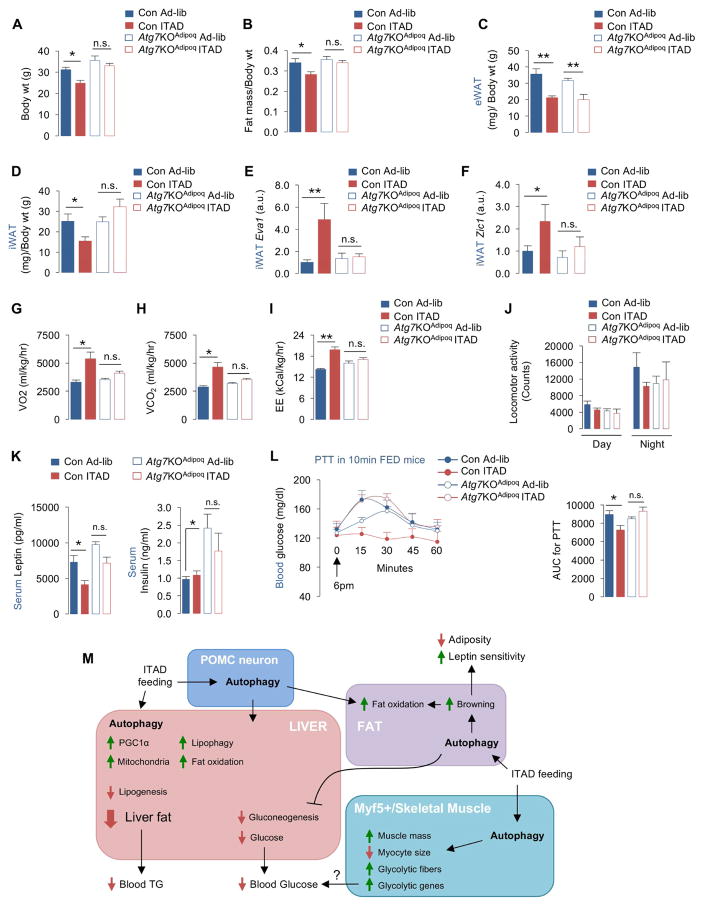
Comment in
-
Metabolism: Metabolic effects of intermeal fasting.Nat Rev Endocrinol. 2018 Jan;14(1):4. doi: 10.1038/nrendo.2017.153. Epub 2017 Nov 10. Nat Rev Endocrinol. 2018. PMID: 29125143 No abstract available.
References
-
- Anson RM, Guo Z, de Cabo R, Iyun T, Rios M, Hagepanos A, Ingram DK, Lane MA, Mattson MP. Intermittent fasting dissociates beneficial effects of dietary restriction on glucose metabolism and neuronal resistance to injury from calorie intake. Proceedings of the National Academy of Sciences of the United States of America. 2003;100:6216–6220. - PMC - PubMed
-
- Badman MK, Pissios P, Kennedy AR, Koukos G, Flier JS, Maratos-Flier E. Hepatic fibroblast growth factor 21 is regulated by PPARalpha and is a key mediator of hepatic lipid metabolism in ketotic states. Cell metabolism. 2007;5:426–437. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
