

Dietary Restriction and AMPK Increase Lifespan via Mitochondrial Network and Peroxisome Remodeling
- PMID: 29107506
- PMCID: PMC5718936
- DOI: 10.1016/j.cmet.2017.09.024
Dietary Restriction and AMPK Increase Lifespan via Mitochondrial Network and Peroxisome Remodeling
Abstract
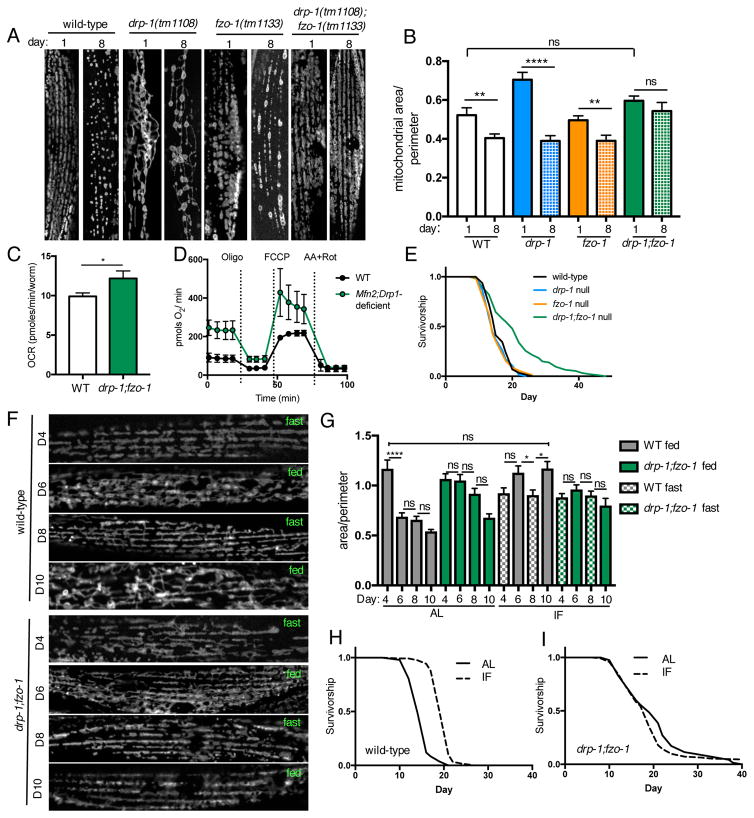
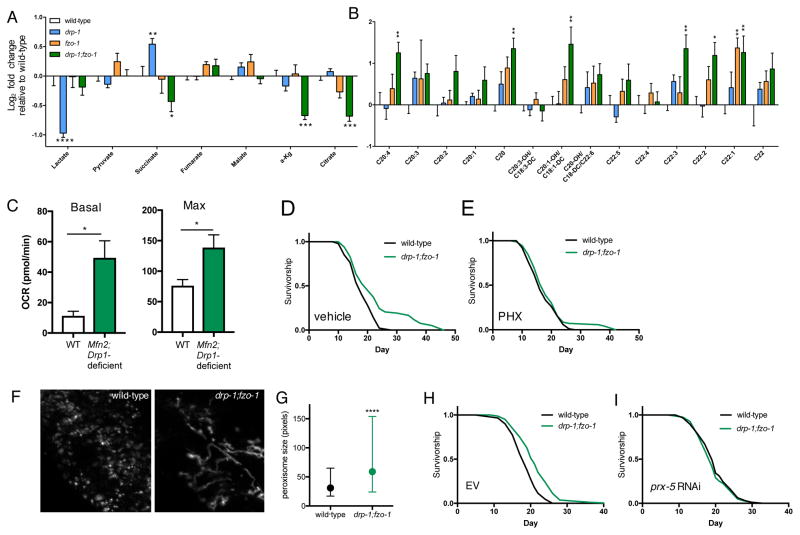
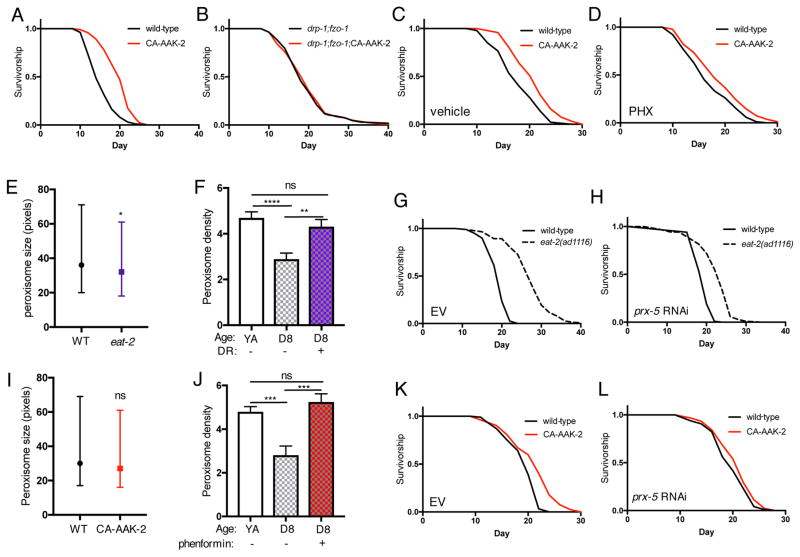
Mitochondrial network remodeling between fused and fragmented states facilitates mitophagy, interaction with other organelles, and metabolic flexibility. Aging is associated with a loss of mitochondrial network homeostasis, but cellular processes causally linking these changes to organismal senescence remain unclear. Here, we show that AMP-activated protein kinase (AMPK) and dietary restriction (DR) promote longevity in C. elegans via maintaining mitochondrial network homeostasis and functional coordination with peroxisomes to increase fatty acid oxidation (FAO). Inhibiting fusion or fission specifically blocks AMPK- and DR-mediated longevity. Strikingly, however, preserving mitochondrial network homeostasis during aging by co-inhibition of fusion and fission is sufficient itself to increase lifespan, while dynamic network remodeling is required for intermittent fasting-mediated longevity. Finally, we show that increasing lifespan via maintaining mitochondrial network homeostasis requires FAO and peroxisomal function. Together, these data demonstrate that mechanisms that promote mitochondrial homeostasis and plasticity can be targeted to promote healthy aging.
Keywords: AMPK; aging; dietary restriction; fatty acid oxidation; intermittent fasting; longevity; mitochondrial dynamics; peroxisomes.
Copyright © 2017 Elsevier Inc. All rights reserved.
Figures
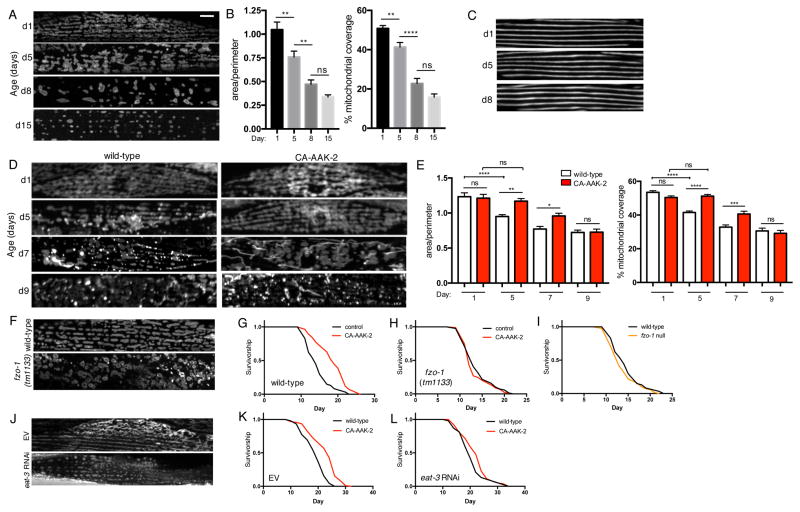
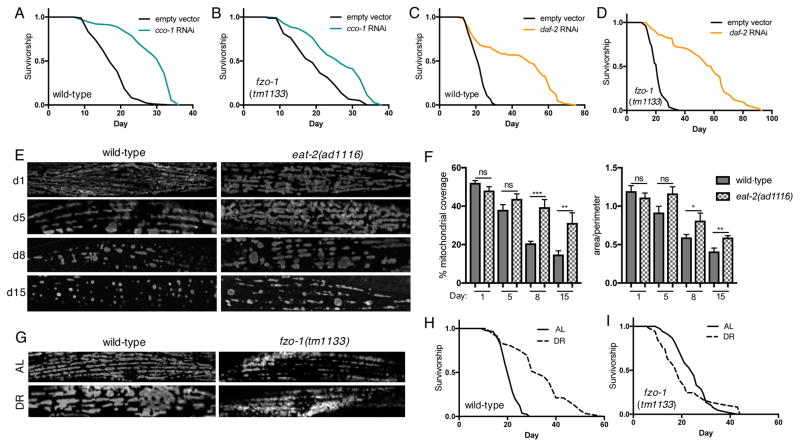
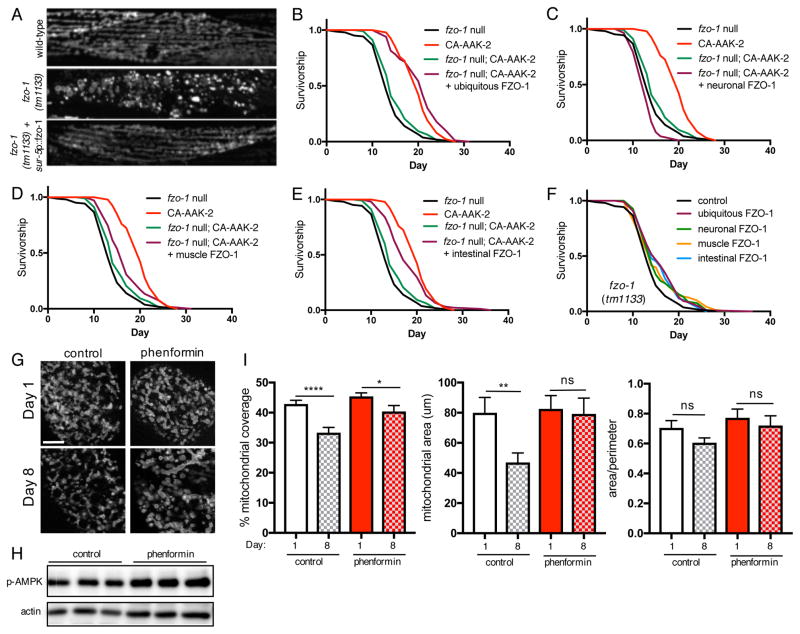
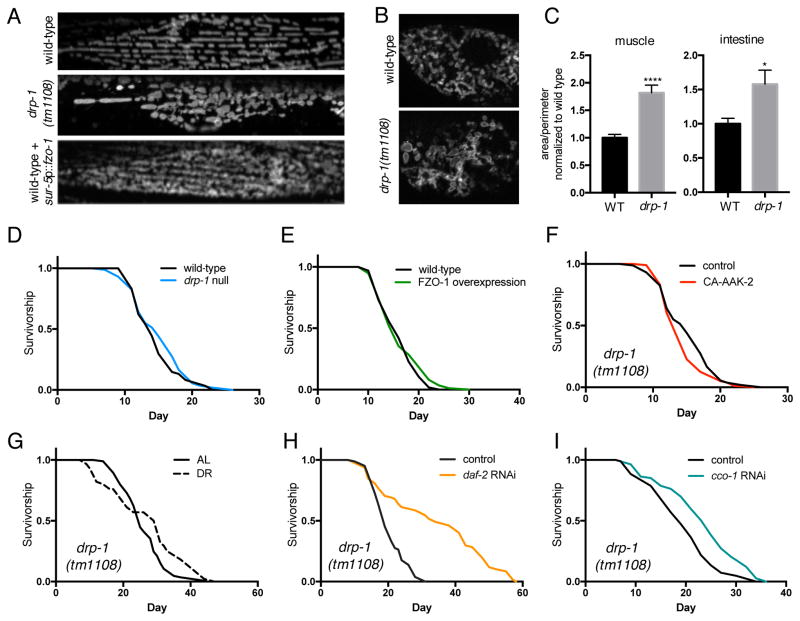
Comment in
-
RevAMPing Mitochondrial Shape to Live Longer.Cell Metab. 2017 Dec 5;26(6):805-806. doi: 10.1016/j.cmet.2017.11.010. Cell Metab. 2017. PMID: 29211978
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
