

Syntenin mediates SRC function in exosomal cell-to-cell communication
- PMID: 29109268
- PMCID: PMC5703317
- DOI: 10.1073/pnas.1713433114
Syntenin mediates SRC function in exosomal cell-to-cell communication
Abstract
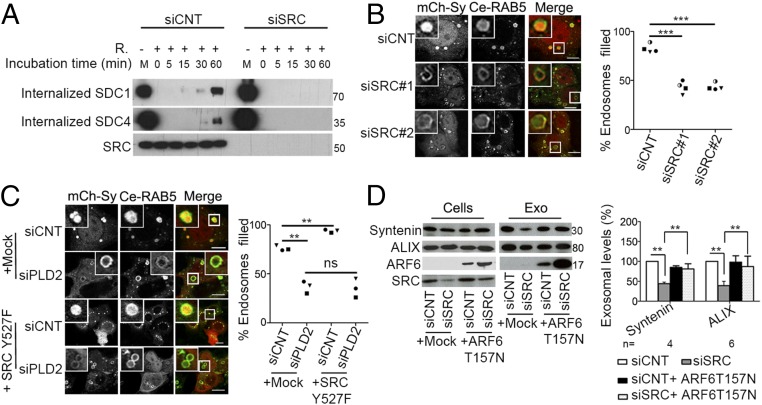
The cytoplasmic tyrosine kinase SRC controls cell growth, proliferation, adhesion, and motility. The current view is that SRC acts primarily downstream of cell-surface receptors to control intracellular signaling cascades. Here we reveal that SRC functions in cell-to-cell communication by controlling the biogenesis and the activity of exosomes. Exosomes are viral-like particles from endosomal origin that can reprogram recipient cells. By gain- and loss-of-function studies, we establish that SRC stimulates the secretion of exosomes having promigratory activity on endothelial cells and that syntenin is mandatory for SRC exosomal function. Mechanistically, SRC impacts on syndecan endocytosis and on syntenin-syndecan endosomal budding, upstream of ARF6 small GTPase and its effector phospholipase D2, directly phosphorylating the conserved juxtamembrane DEGSY motif of the syndecan cytosolic domain and syntenin tyrosine 46. Our study uncovers a function of SRC in cell-cell communication, supported by syntenin exosomes, which is likely to contribute to tumor-host interactions.
Keywords: ARF6; SRC; exosome; syndecan; syntenin.
Copyright © 2017 the Author(s). Published by PNAS.
Conflict of interest statement
The authors declare no conflict of interest.
Figures




References
-
- Yeatman TJ. A renaissance for SRC. Nat Rev Cancer. 2004;4:470–480. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
