

Oxytocin Removes Estrous Female vs. Male Preference of Virgin Male Rats: Mediation of the Supraoptic Nucleus Via Olfactory Bulbs
- PMID: 29109676
- PMCID: PMC5660071
- DOI: 10.3389/fncel.2017.00327
Oxytocin Removes Estrous Female vs. Male Preference of Virgin Male Rats: Mediation of the Supraoptic Nucleus Via Olfactory Bulbs
Abstract
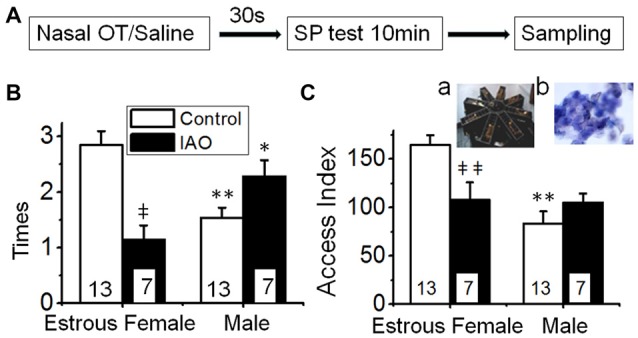
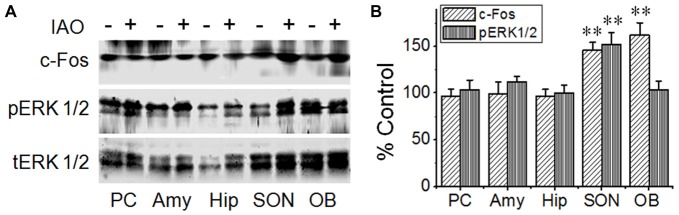
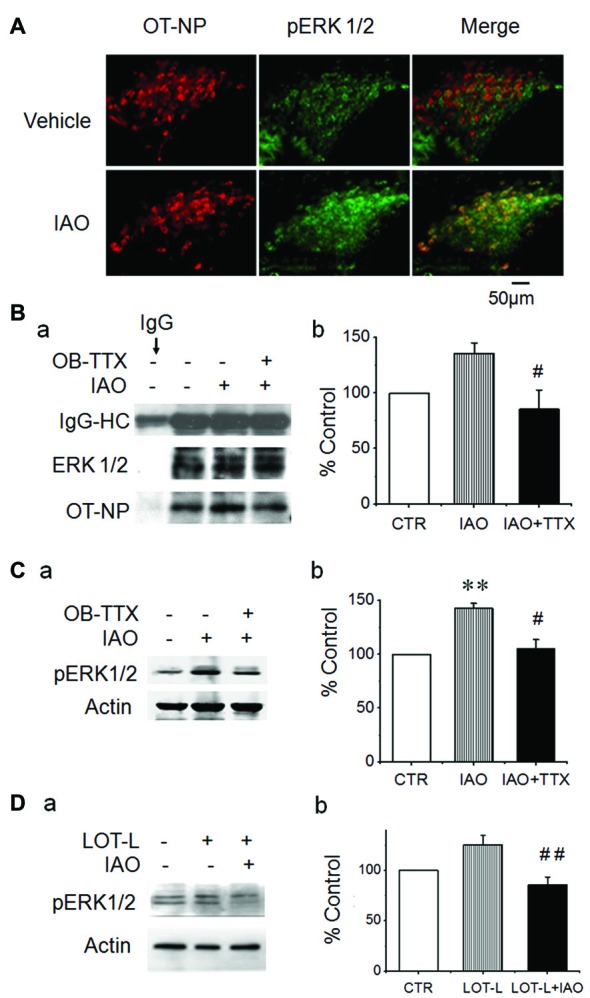
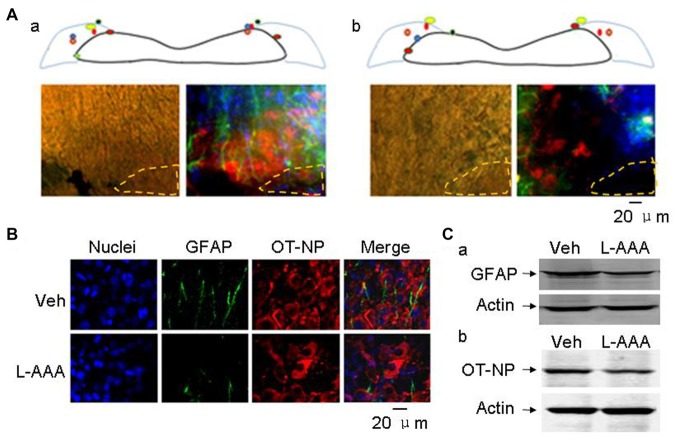
Social functions of oxytocin (OT) have been explored extensively; however, relationship between the effect of intranasally applied OT (nasal OT) on the social preference (SP) and intracerebral actions of endogenous OT remains unclear. To resolve this question, we first observed effects of nasal OT on the SP of virgin young adult male rats toward unfamiliar virgin estrous female (EF) vs. virgin male rats. The results showed that the test male rats exhibited significantly more times and longer duration accessing the female than the male, which were acutely eliminated by nasal OT. Then, we examined the approaches mediating nasal OT effects on the activity of potential brain targets in Western blots and found that nasal OT activated the olfactory bulbs (OBs) and the supraoptic nucleus (SON), but not the piriform cortex, amygdala and hippocampus as shown by significant changes in the expression of c-Fos and/or phosphorylated extracellular signal-regulated protein kinase (pERK) 1/2. Moreover, microinjection of TTX into the OBs blocked nasal OT-evoked increases in pERK1/2 levels as well as the molecular association between ERK1/2 and OT-neurophysin in the SON. Electrolytic lesions of the lateral olfactory tract did not significantly change the basal levels of pERK 1/2 in the SON; however, upon nasal OT, pERK 1/2 levels in the SON reduced significantly. Lastly, microinjection of L-aminoadipic acid (gliotoxin) into the SON to reduce OT levels reduced the duration of the test male's accessing the EF and blocked the nasal OT-evoked increase in the duration of test male's accessing the male while significantly increasing pERK1/2 levels in the amygdala. These findings reveal for the first time that nasal OT acutely eliminates virgin males' SP to EFs via the OB-SON route and that OT neurons could mediate the social effects of nasal OT by suppressing social phobia generated in the amygdala.
Keywords: intranasal drug delivery; olfactory bulbs; oxytocin; social preference; supraoptic nucleus.
Figures
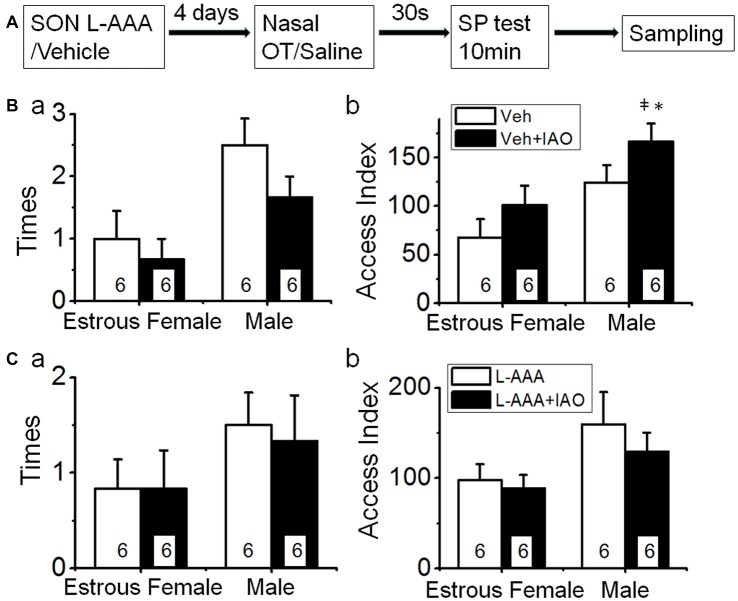
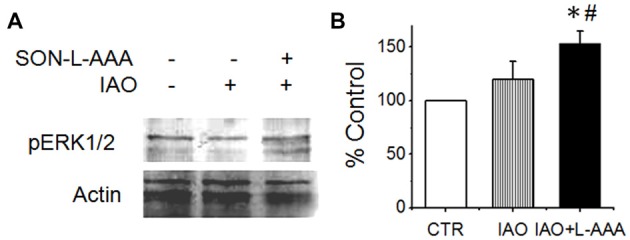
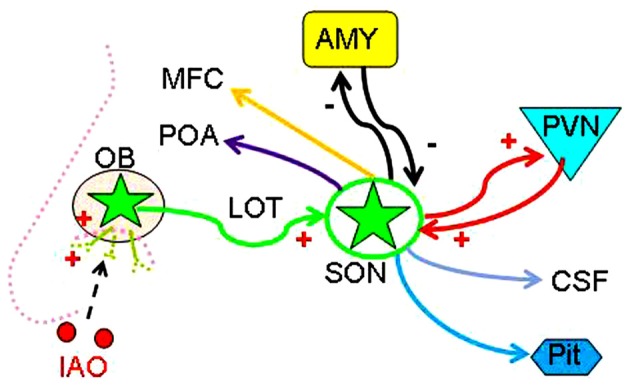
References
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous