

Targeting de novo lipogenesis as a novel approach in anti-cancer therapy
- PMID: 29112683
- PMCID: PMC5765225
- DOI: 10.1038/bjc.2017.374
Targeting de novo lipogenesis as a novel approach in anti-cancer therapy
Abstract
Background: Although altered membrane physiology has been discussed within the context of cancer, targeting membrane characteristics by drugs being an attractive therapeutic strategy has received little attention so far.
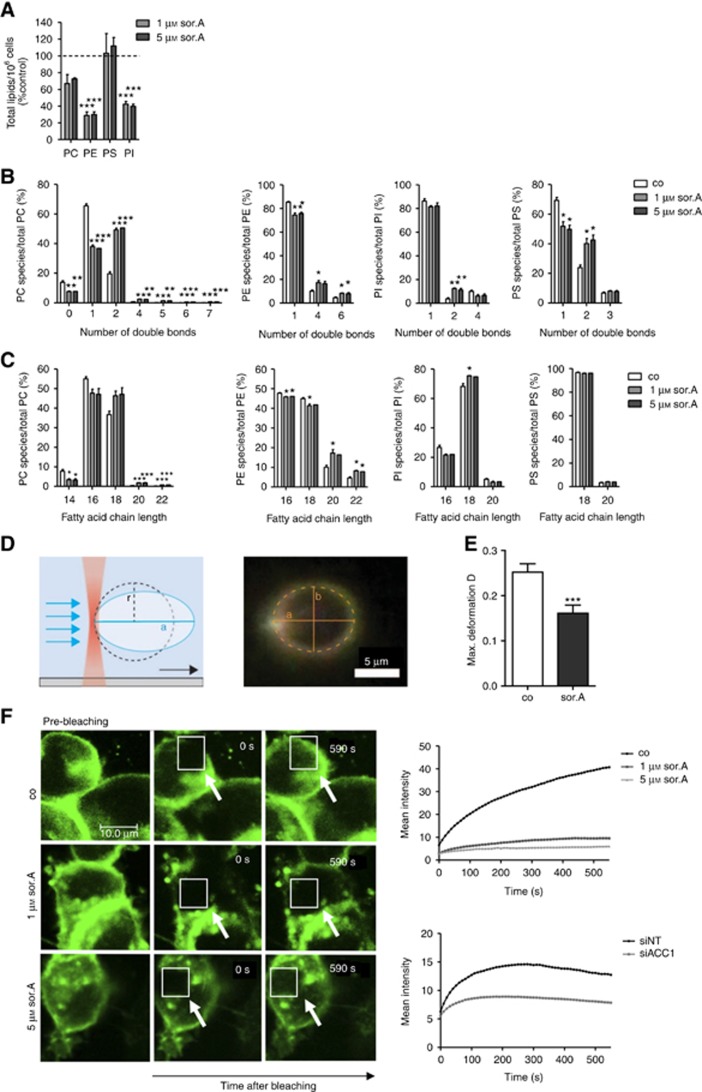
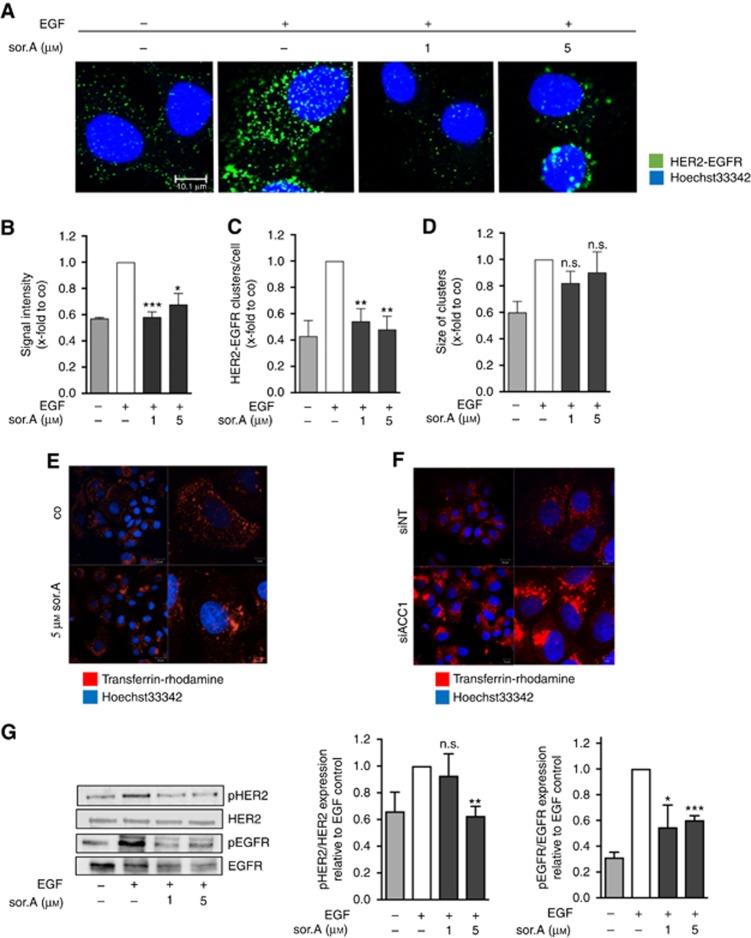
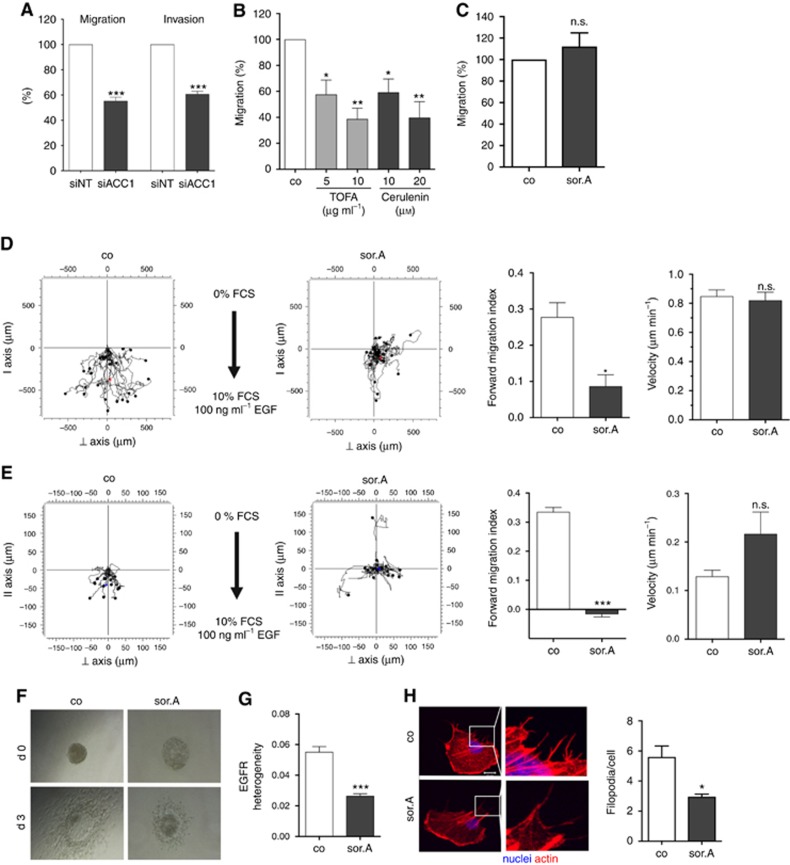
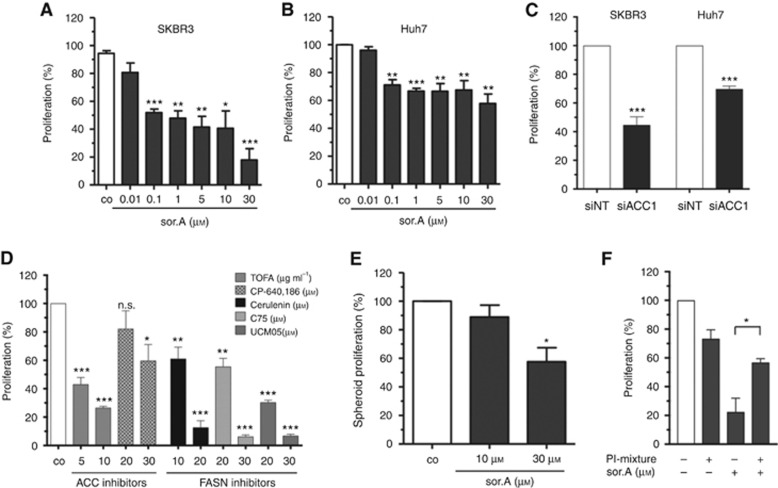
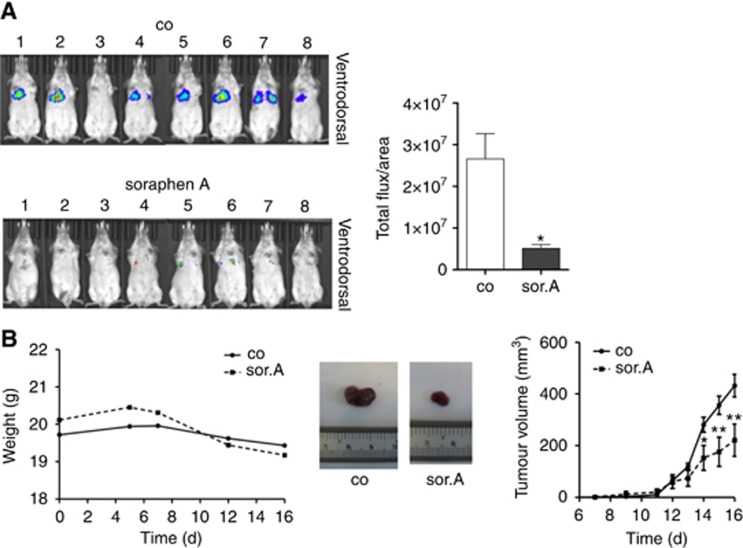
Methods: Various acetyl-CoA carboxylase 1 (ACC1), and fatty acid synthase (FASN) inhibitors (like Soraphen A and Cerulenin) as well as genetic knockdown approaches were employed to study the effects of disturbed phospholipid composition on membrane properties and its functional impact on cancer progression. By using state-of-the-art methodologies such as LC-MS/MS, optical tweezers measurements of giant plasma membrane vesicles and fluorescence recovery after photobleaching analysis, membrane characteristics were examined. Confocal laser scanning microscopy, proximity ligation assays, immunoblotting as well as migration, invasion and proliferation experiments unravelled the functional relevance of membrane properties in vitro and in vivo.
Results: By disturbing the deformability and lateral fluidity of cellular membranes, the dimerisation, localisation and recycling of cancer-relevant transmembrane receptors is compromised. Consequently, impaired activation of growth factor receptor signalling cascades results in abrogated tumour growth and metastasis in different in vitro and in vivo models.
Conclusions: This study highlights the field of membrane properties as a promising druggable cellular target representing an innovative strategy for development of anti-cancer agents.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Beckers A, Organe S, Timmermans L, Scheys K, Peeters A, Brusselmans K, Verhoeven G, Swinnen JV (2007) Chemical inhibition of acetyl-CoA carboxylase induces growth arrest and cytotoxicity selectively in cancer cells. Cancer Res 67: 8180–8187. - PubMed
-
- Berod L, Friedrich C, Nandan A, Freitag J, Hagemann S, Harmrolfs K, Sandouk A, Hesse C, Castro CN, Bahre H, Tschirner SK, Gorinski N, Gohmert M, Mayer CT, Huehn J, Ponimaskin E, Abraham WR, Muller R, Lochner M, Sparwasser T (2014) De novo fatty acid synthesis controls the fate between regulatory T and T helper 17 cells. Nat Med 20: 1327–1333. - PubMed
-
- Braig S, Schmidt BUS, Katharina S, Chris H, Till M, Oliver W, Rolf M, Stefan Z, Andreas K, Josef AK, Angelika MV (2015) Pharmacological targeting of membrane rigidity: implications on cancer cell migration and invasion. N J Phys 17: 083007.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
