

G-quadruplex-binding small molecules ameliorate C9orf72 FTD/ALS pathology in vitro and in vivo
- PMID: 29113975
- PMCID: PMC5760849
- DOI: 10.15252/emmm.201707850
G-quadruplex-binding small molecules ameliorate C9orf72 FTD/ALS pathology in vitro and in vivo
Abstract
Intronic GGGGCC repeat expansions in C9orf72 are the most common known cause of frontotemporal dementia (FTD) and amyotrophic lateral sclerosis (ALS), which are characterised by degeneration of cortical and motor neurons, respectively. Repeat expansions have been proposed to cause disease by both the repeat RNA forming foci that sequester RNA-binding proteins and through toxic dipeptide repeat proteins generated by repeat-associated non-ATG translation. GGGGCC repeat RNA folds into a G-quadruplex secondary structure, and we investigated whether targeting this structure is a potential therapeutic strategy. We performed a screen that identified three structurally related small molecules that specifically stabilise GGGGCC repeat G-quadruplex RNA We investigated their effect in C9orf72 patient iPSC-derived motor and cortical neurons and show that they significantly reduce RNA foci burden and the levels of dipeptide repeat proteins. Furthermore, they also reduce dipeptide repeat proteins and improve survival in vivo, in GGGGCC repeat-expressing Drosophila Therefore, small molecules that target GGGGCC repeat G-quadruplexes can ameliorate the two key pathologies associated with C9orf72 FTD/ALS These data provide proof of principle that targeting GGGGCC repeat G-quadruplexes has therapeutic potential.
Keywords: C9orf72; G‐quadruplex; amyotrophic lateral sclerosis; frontotemporal dementia.
© 2017 The Authors. Published under the terms of the CC BY 4.0 license.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures

A FRET assay was used to measure the difference in melting temperature (ΔTm) of (G4C2)4 RNA or DNA G‐Qs in the presence of 2 μM of 138 different small molecules. An increase in ΔTm indicates stabilisation of the G‐Q. Small molecules are ranked on the x‐axis according to their increasing thermal stabilisation of the DNA (G4C2)4 G‐Q. Small molecules that preferentially stabilise RNA over DNA (G4C2)4 G‐Qs reside in the upper part of the scatter plot, above the blue curve. An arbitrary ΔTm threshold of 13°C greater than vehicle (grey line) and a differential binding to RNA over DNA (ΔTmRNA–ΔTmDNA ≥ 5°C) were used to select candidate small molecules.
Structures of the three compounds (DB1246, DB1247, DB1273), highlighted by green circles in (A), that show preferential binding to RNA (G4C2)4 G‐Qs and were further characterised.
FRET dose response of DB1246, DB1247 and DB1273 on stabilisation of RNA or DNA (G4C2)4 G‐Qs.
Temperature unfold CD spectra for (G4C2)4 RNA alone (which shows a characteristic G‐Q structure with minima at 237 nm, maxima at 264 nm and no additional signal), or in the presence of 2 μM DB1246, DB1247 or DB1273. A characteristic induced CD spectrum, in the 350–550 nm region, is observed only in the presence of each small molecule, confirming that each of these three compounds are binding to (G4C2)4 RNA G‐Qs.

C9orf72 iPSC‐motor neurons express choline acetyltransferase (ChAT) and beta‐tubulin (TUJ1). Scale bar represents 10 μm.
The percentage of total cells positive for ChAT was quantified after differentiation into motor neurons. No difference was observed between control and C9orf72‐derived iPSC‐motor neurons, with approximately 90% of all cells converted to ChAT‐positive motor neurons. Two independent control iPSC lines and three independent C9orf72 lines were analysed, with one to two independent differentiations per line and > 100 cells quantified per line. Bars show the average and SEM. P > 0.05, Mann–Whitney U‐test.

Representative images of RNA foci (red) within iPSC‐cortical neurons; nuclei are visualised with DAPI (blue). Scale bar represents 10 μm.
Quantification shows RNA foci are significantly reduced by all three small molecules, DB1246, DB1247 and DB1273, at a concentration of 1 μM for 4 days. Data are shown as the average and SD of the percentage of neurons containing RNA foci in 5–10 40× fields of view for one C9orf72 patient iPSC‐cortical neuron line. ***P = 0.0006 (DB1246, 0.5 μM), ***P = 0.0001 (DB1247, 1 μM), ***P = 0.0001 (DB1273, 1 μM), one‐way ANOVA with Dunnett's post hoc test versus DMSO.

Representative images of C9orf72 iPSC‐cortical neurons treated with DMSO (vehicle control) or 1 μM of DB1246, DB1247 or DB1273, for 4 days. RNA foci are shown in red and nuclei (DAPI) in blue. Scale bar represents 10 μm.
Quantification shows RNA foci are significantly reduced by all three compounds in iPSC‐cortical neurons and by DB1246 and DB1273 in iPSC‐motor neurons. Data are shown as the average percentage of neurons containing RNA foci relative to vehicle (DMSO). N = 3 independent C9orf72 patient lines with two to three inductions per line and at least 70 neurons counted per induction, data are shown as mean and SEM. *P < 0.05, **P < 0.01, ***P < 0.001, one‐sample two‐tailed t‐test versus normalised control. For cortical neurons, *P = 0.0124 (DB1246), **P = 0.0065 (DB1247), **P = 0.0096 (DB1273). For motor neurons, ***P = 0.0004 (DB1246), **P = 0.0030 (DB1273).

Poly(GP) levels were measured by MSD immunoassay in C9orf72 patient iPSC‐motor neurons treated for 7 days with 8–16 μM of the G4C2 repeat G‐Q‐binding small molecules DB1246, DB1247 and DB1273. Treatment with DB1246 or DB1273 leads to a significant reduction in poly(GP) levels relative to vehicle‐treated controls. Data are shown as the mean and SEM of three independent C9orf72 iPSC lines, with one to six differentiations per line. **P = 0.0068 (DB1246, 12 μm), *P = 0.0417 (DB1246, 16 μm), *P = 0.0196 (DB1247, 8 μm), ***P = 0.0002 (DB1273, 8 μm), *P = 0.0194 (DB1273, 12 μm), **P = 0.0062 (DB1273, 16 μm), one‐sample two‐tailed t‐test versus normalised control.
C9orf72 patient iPSC‐motor neurons were treated for 7 days with 16 μM of each small molecule and the expression levels of MCM2 and the three C9orf72 transcript variants (V1–V3) measured by quantitative RT–PCR. Data are shown as the mean and SD of three independent iPSC‐motor neuron lines (one induction per line) relative to vehicle (DMSO)‐treated controls. No significant changes in gene expression were observed, one‐way ANOVA, Dunnett's post hoc test, P > 0.05.
XTT cell death assay for C9orf72 iPSC‐motor neurons treated for 7 days with 0–40 μM of DB1246, DB1247, DB1273 or cisplatin as a positive control. Data are shown as the mean and SEM of two independent iPSC‐motor neuron lines. Toxicity is only observed at the highest dose of 40 μM and not at the penultimate dose of 20 μM.
Drosophila first‐instar larvae expressing 36 G4C2 repeats were placed on food containing either vehicle or 1 mM DB1273, and the number reaching the pupal stage of development counted after 7 days. DB1273 treatment (n = 1,200 flies) significantly improves survival compared to vehicle (n = 1,150 flies), *P = 0.0320, chi‐squared test. Data are shown as proportion reaching the pupal stage, with numbers within each group indicated on the bars. Genotype was w1118; UAS‐36(GGGGCC)/+; daGAL4/+ (daGAL4>36R).
Drosophila first‐instar larvae ubiquitously expressing 36 G4C2 repeats (36R) were treated with vehicle or 1 mM DB1273 for 5 days, until the third‐instar stage (L3), and poly(GR) measured by MSD immunoassay. Poly(GR) was also measured in control (w1118) larvae that do not express G4C2 repeats. DB1273 treatment significantly reduced poly(GR) expression, data are shown as mean ± SEM. n = 8 for 36R groups, n = 7 for control group. ***P = 0.0001 (w1118 versus 36R 0 mM larvae), **P = 0.0019 (0 mM versus 1 mM 36R larvae), one‐way ANOVA with Dunnett's post hoc test. Genotypes were w1118; and w1118; UAS‐36(GGGGCC)/+; daGAL4/+ (daGAL4>36R).
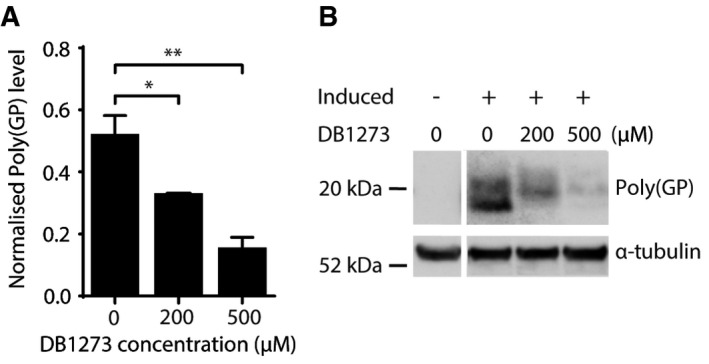
Treatment of adult Drosophila ubiquitously expressing 36 G4C2 repeats for 7 days with DB1273 leads to a significant reduction in poly(GP) levels (normalised to α‐tubulin) relative to vehicle‐treated controls. Data are shown as the mean and SEM of three experiments. *P = 0.0463 (200 μM versus 0 μM), **P = 0.0026 (500 μM versus 0 μM), one‐way ANOVA with Dunnett's post hoc test. Genotype was w1118; daGS/UAS‐36(GGGGCC) (daGS>36R).
Representative poly(GP) immunoblot. Poly(GP) appears as a doublet in the induced daGS>36R flies and is not present in uninduced flies, confirming its generation is due to expression of the (G4C2)36 repeats. Genotype was w1118; daGS/UAS‐36(GGGGCC) (daGS>36R).

The MSD immunoassay specifically detects (GR)7 peptide but not other dipeptide repeat proteins (GP)7 or (PR)7.
Representative standard curve of MSD response at different concentrations of (GR)7 peptide calibrator, R 2 = 0.99.
Enhanced view of (B) to show the lower limit of detection of the assay, which is 0.125 ng/ml (dashed line).

Comment in
-
Targeting RNA G-quadruplexes as new treatment strategy for C9orf72 ALS/FTD.EMBO Mol Med. 2018 Jan;10(1):4-6. doi: 10.15252/emmm.201708572. EMBO Mol Med. 2018. PMID: 29175945 Free PMC article.
References
-
- Beck J, Poulter M, Hensman D, Rohrer JD, Mahoney CJ, Adamson G, Campbell T, Uphill J, Borg A, Fratta P et al (2013) Large C9orf72 hexanucleotide repeat expansions are seen in multiple neurodegenerative syndromes and are more frequent than expected in the UK population. Am J Hum Genet 92: 345–353 - PMC - PubMed
-
- van Blitterswijk M, Dejesus‐Hernandez M, Niemantsverdriet E, Murray ME, Heckman MG, Diehl NN, Brown PH, Baker MC, Finch NA, Bauer PO et al (2013) Association between repeat sizes and clinical and pathological characteristics in carriers of C9ORF72 repeat expansions (Xpansize‐72): a cross‐sectional cohort study. Lancet Neurol 12: 978–988 - PMC - PubMed
Publication types
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
