

Response to Trypanosoma cruzi by Human Blood Cells Enriched with Dentritic Cells Is Controlled by Cyclooxygenase-2 Pathway
- PMID: 29118740
- PMCID: PMC5660982
- DOI: 10.3389/fmicb.2017.02020
Response to Trypanosoma cruzi by Human Blood Cells Enriched with Dentritic Cells Is Controlled by Cyclooxygenase-2 Pathway
Abstract
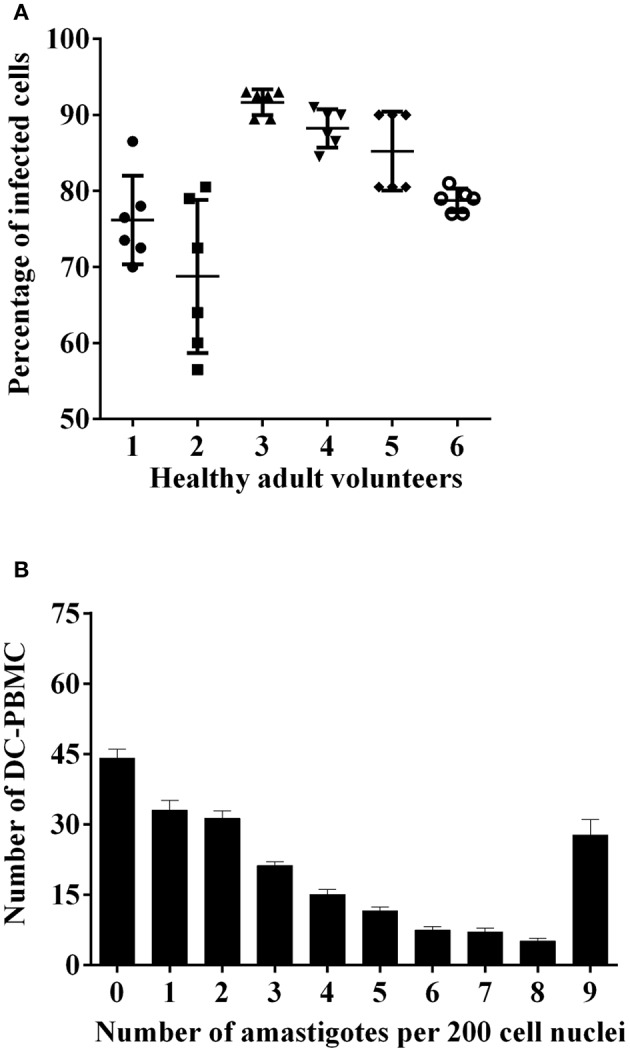
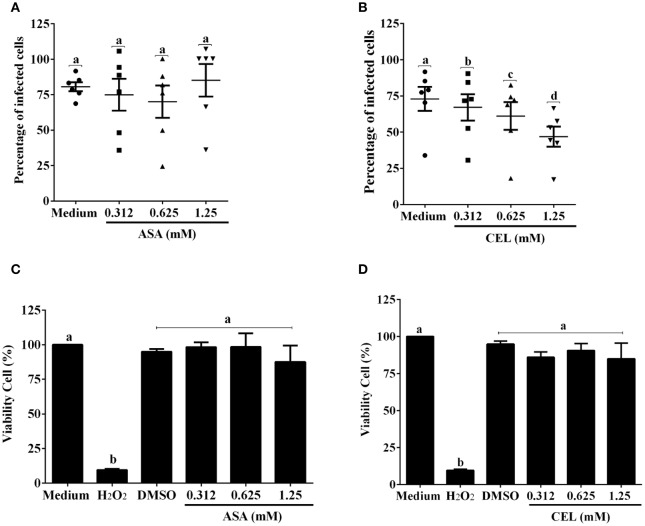
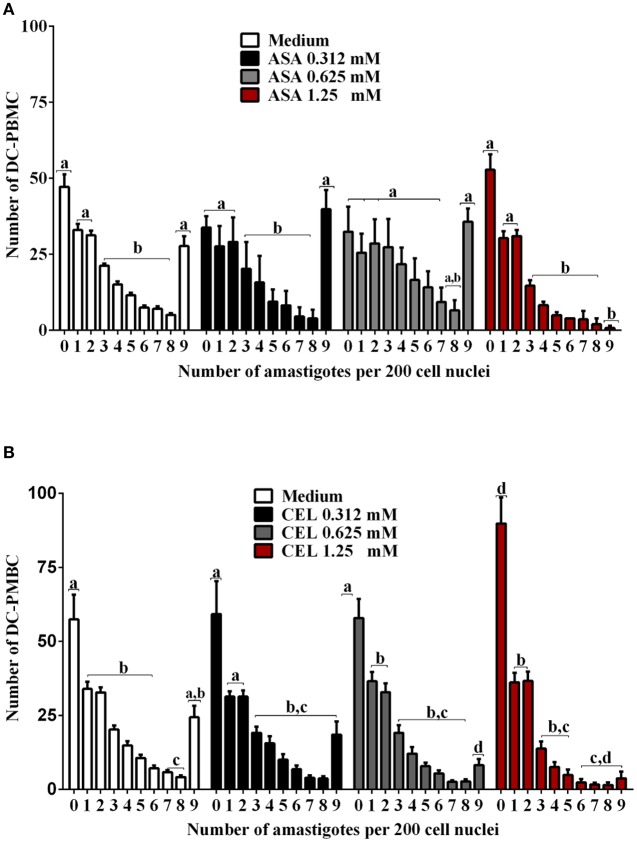
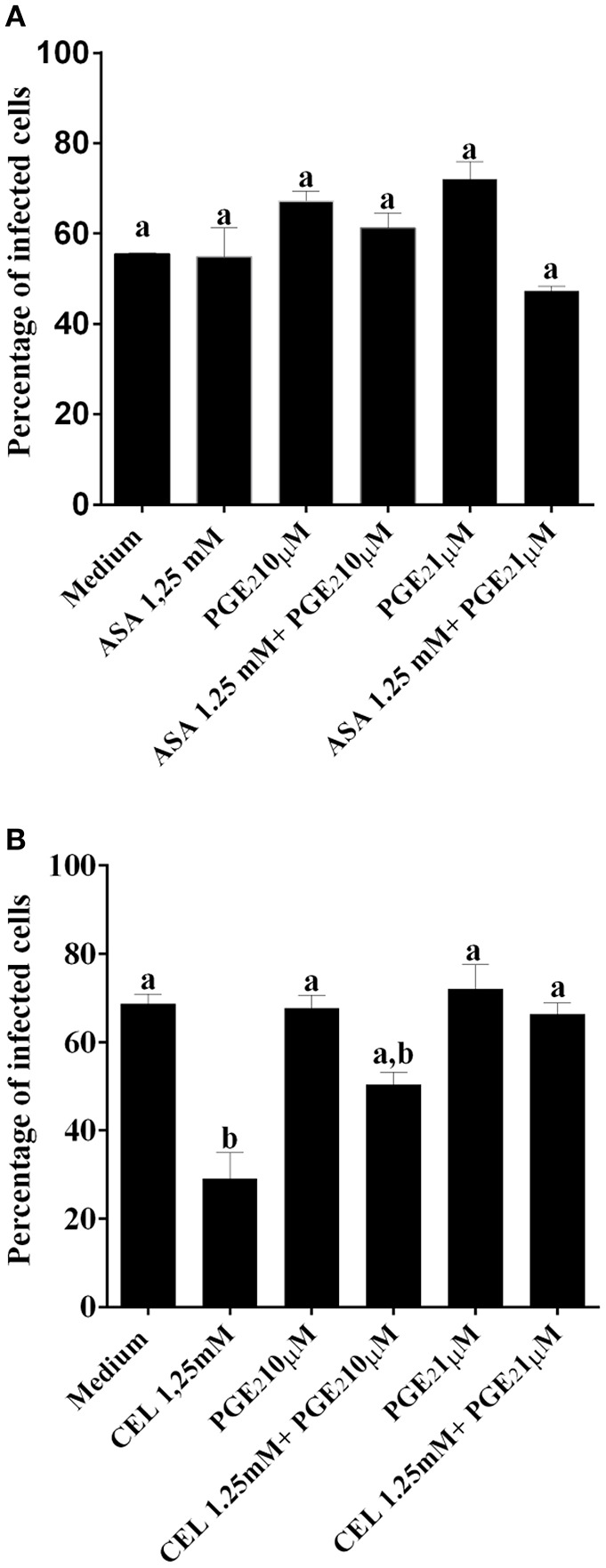
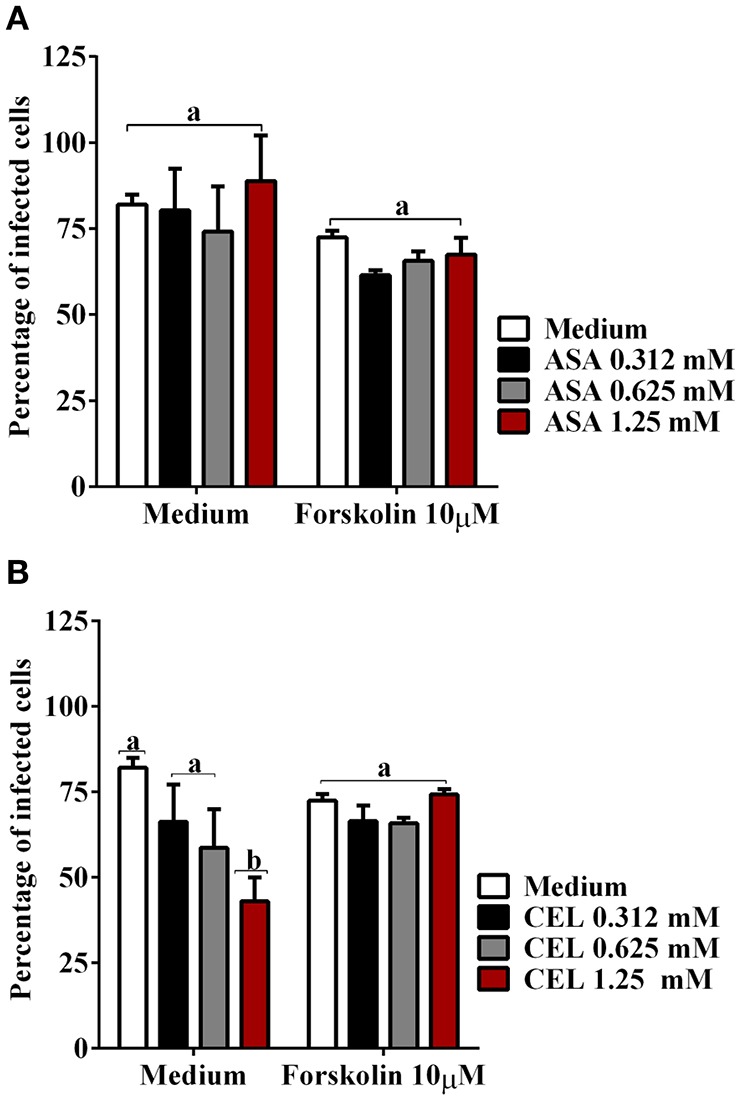
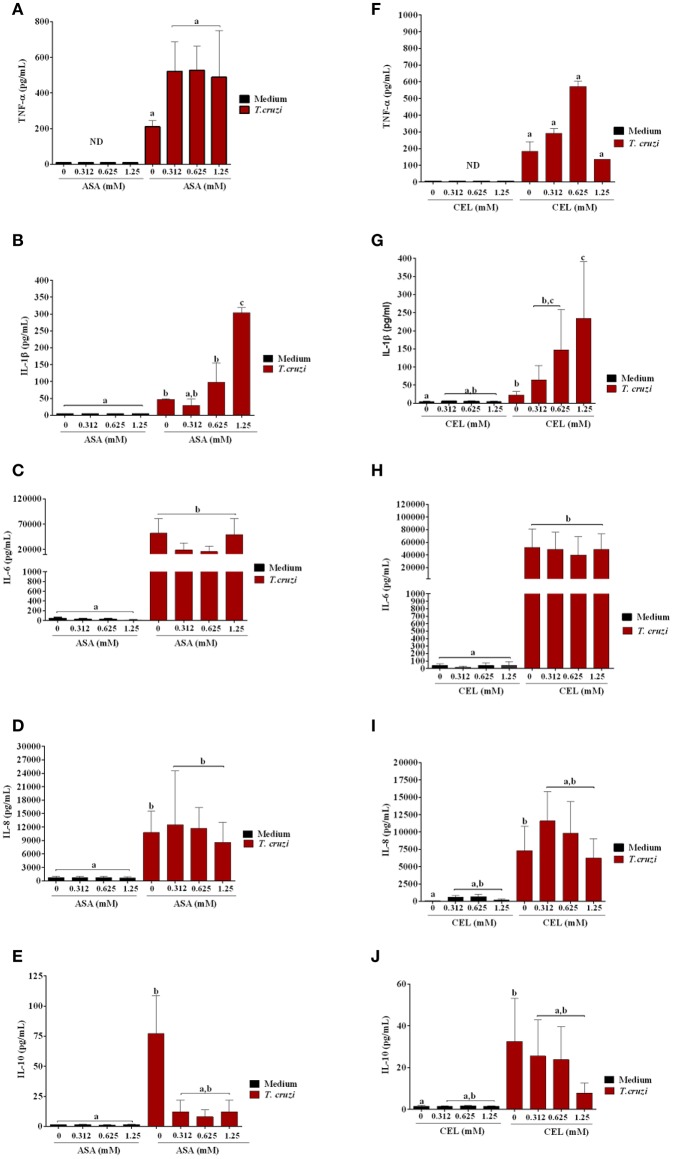
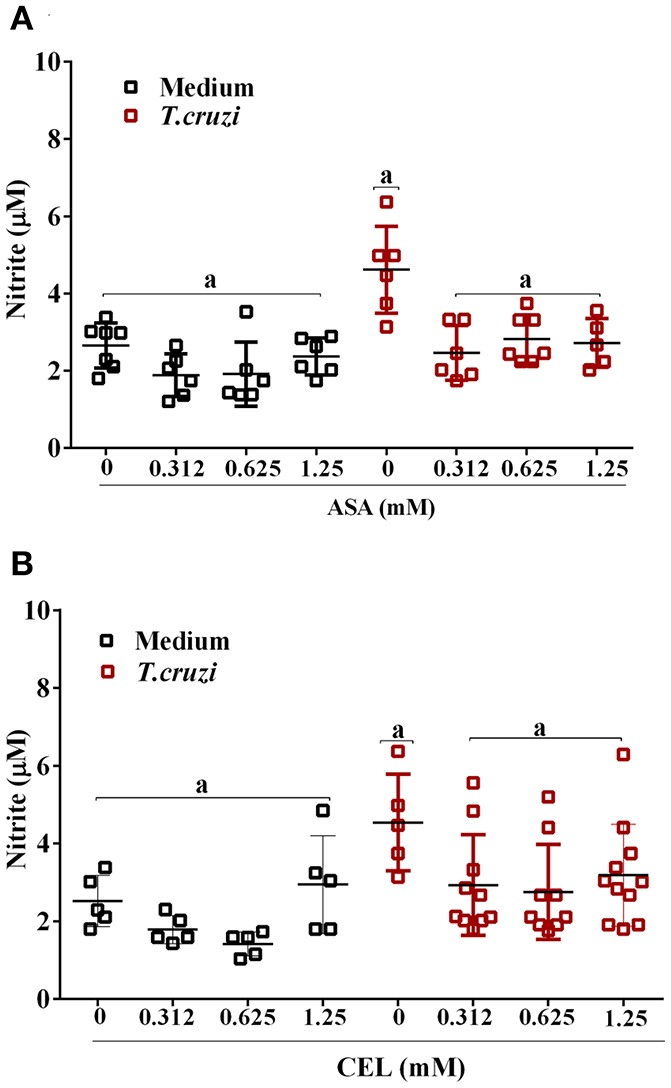
Chagas disease (Cd) or American human trypanosomiasis is caused by Trypanosoma cruzi and affects ~7 million people, mostly in Latin America. The infective trypomastigote forms of the parasite can invade several human blood cell populations, including monocytes and dendritic cells (DC). Although these cells display a wide functional diversity, their interactions with T. cruzi via cyclooxygenase (COX) and cyclic adenosine monophosphate (cAMP) dependent pathways have not been analyzed. To exploiting this mechanism, DC-enriched peripheral human blood mononuclear cell populations (DC-PBMC) were used as our model. Our results showed that the treatment of these cell populations with celecoxib (CEL), a cyclooxygenase-2 selective inhibitor or SQ 22,536, an adenilate cyclase inhibitor, significantly caused marked inhibition of T. cruzi infection. In contrast, aspirin (ASA, a non-selective COX-1 and COX-2 inhibitor) treatment did not inhibit the infection of the cells by the parasite and was independent of nitric oxide (NO) production. The expression of co-stimulatory molecules CD80 and CD86 were similar on cells treated or not with both COX-inhibitors. The infection stimulated the release of TNF-α, IL-1β, IL-6, IL-8, and IL-10 production by infected cells. Treatment with ASA or CEL did not affect TNF-α, IL-6, IL-8, IL-10, and NO production by infected cells, but increased IL-1β production by them. Our results suggest a key role of COX-2 and cAMP pathways in T. cruzi invasion process of human blood cells and these pathways may represent targets of new therapeutic options for Cd.
Keywords: Trypanosoma cruzi; aspirin; celecoxib; cell invasion; human monocyte-derived dendritic cells.
Figures

References
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials