

Optimal housing temperatures for mice to mimic the thermal environment of humans: An experimental study
- PMID: 29122558
- PMCID: PMC5784327
- DOI: 10.1016/j.molmet.2017.10.009
Optimal housing temperatures for mice to mimic the thermal environment of humans: An experimental study
Abstract
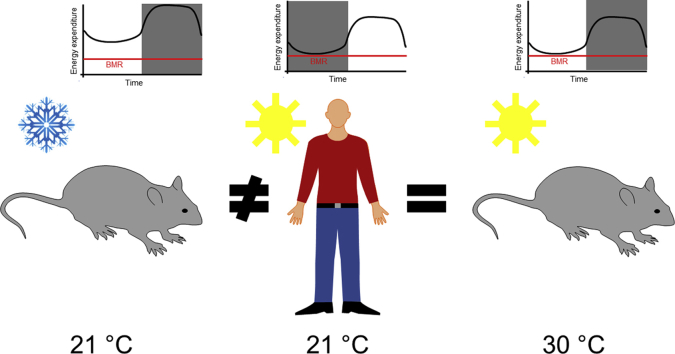
Objectives: The laboratory mouse is presently the most common model for examining mechanisms of human physiology and disease. Housing temperatures can have a large impact on the outcome of such experiments and on their translatability to the human situation. Humans usually create for themselves a thermoneutral environment without cold stress, while laboratory mice under standard conditions (≈20° C) are under constant cold stress. In a well-cited, theoretical paper by Speakman and Keijer in Molecular Metabolism, it was argued that housing mice under close to standard conditions is the optimal way of modeling the human metabolic situation. This tenet was mainly based on the observation that humans usually display average metabolic rates of about 1.6 times basal metabolic rate. The extra heat thereby produced would also be expected to lead to a shift in the 'lower critical temperature' towards lower temperatures.
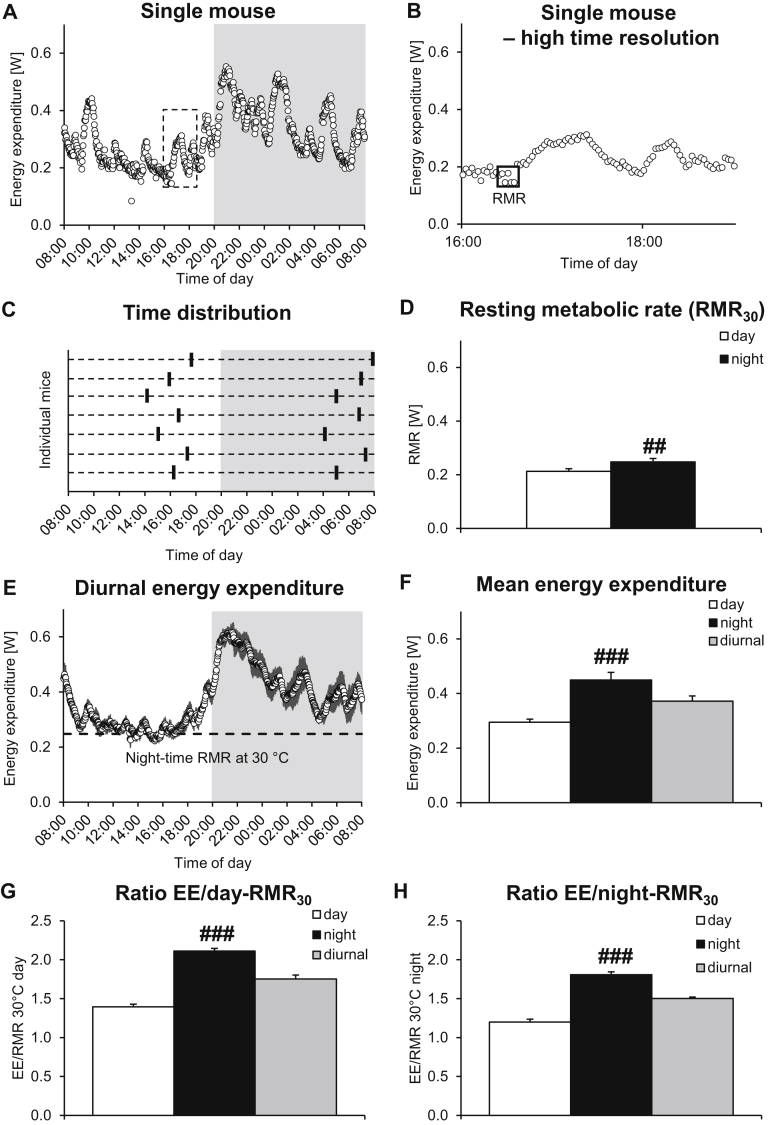
Methods: To examine these tenets experimentally, we performed high time-resolution indirect calorimetry at different environmental temperatures on mice acclimated to different housing temperatures.
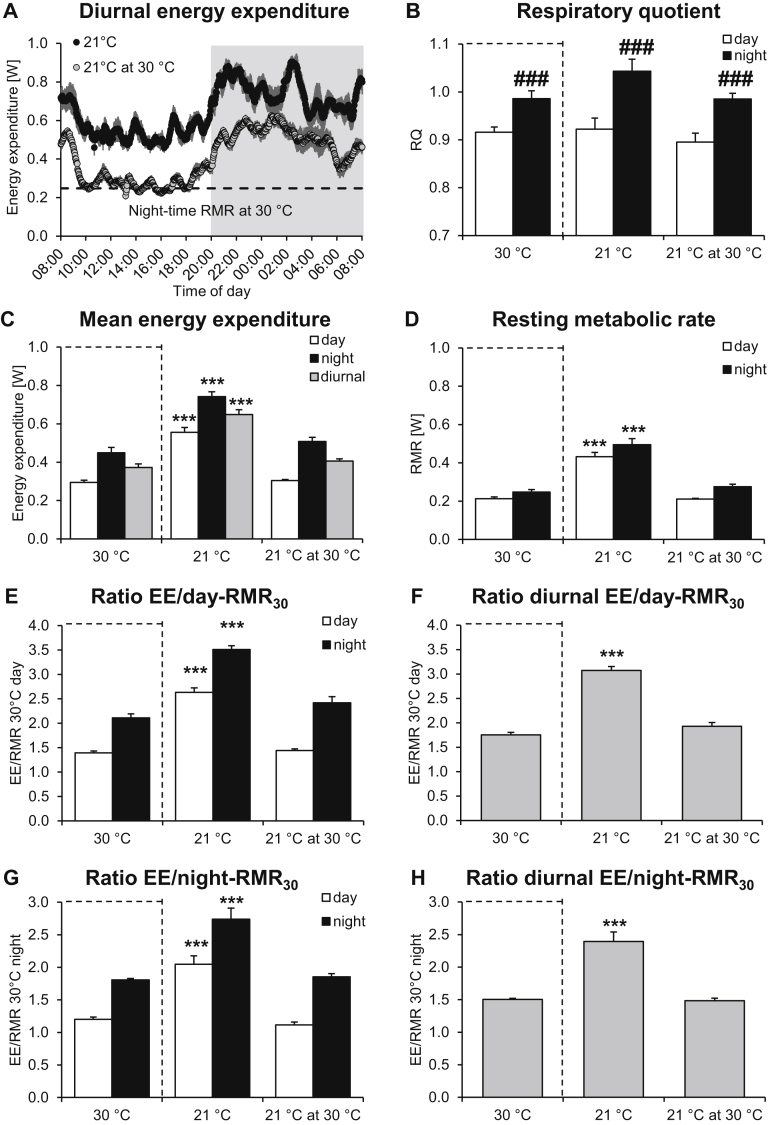
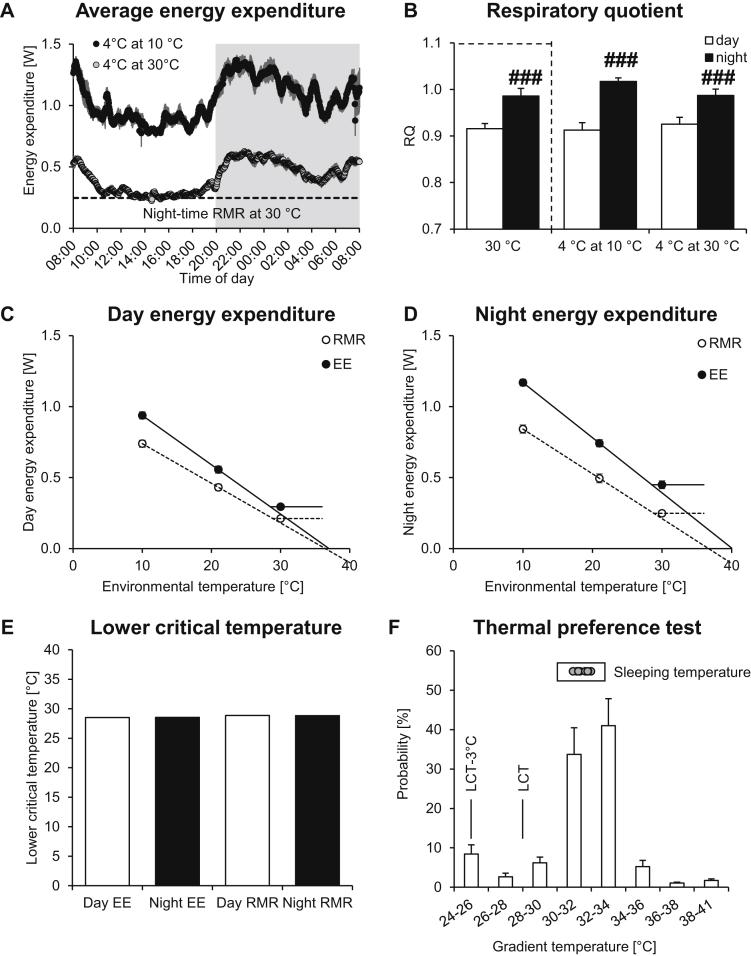
Results: Based on the high time-resolution calorimetry analysis, we found that mice already under thermoneutral conditions display mean diurnal energy expenditure rates 1.8 times higher than basal metabolism, remarkably closely resembling the human situation. At any temperature below thermoneutrality, mice metabolism therefore exceeds the human equivalent: Mice under standard conditions display energy expenditure 3.1 times basal metabolism. The discrepancy to previous conclusions is probably attributable to earlier limitations in establishing true mouse basal metabolic rate, due to low time resolution. We also found that the fact that mean energy expenditure exceeds resting metabolic rate does not move the apparent thermoneutral zone (the lower critical temperature) downwards.
Conclusions: We show that housing mice at thermoneutrality is an advantageous step towards aligning mouse energy metabolism to human energy metabolism.
Keywords: Ambient temperature; Basal metabolic rate; Human; Lower critical temperature; Mouse; Thermoneutral; Thermoregulation.
Copyright © 2017 The Authors. Published by Elsevier GmbH.. All rights reserved.
Figures
References
-
- Abreu-Vieira G., Fischer A.W., Mattsson C., de Jong J.M.A., Shabalina I.G., Rydén M. Cidea improves the metabolic profile through expansion of adipose tissue. Nature Communications. 2015;6:7433. - PubMed
-
- Cannon B., Nedergaard J. Nonshivering thermogenesis and its adequate measurement in metabolic studies. Journal of Experimental Biology. 2011;214:242–253. - PubMed
-
- Feldmann H.M., Golozoubova V., Cannon B., Nedergaard J. UCP1 ablation induces obesity and abolishes diet-induced thermogenesis in mice exempt from thermal stress by living at thermoneutrality. Cell Metabolism. 2009;9:203–209. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
