

Genome-Wide Screen for Saccharomyces cerevisiae Genes Contributing to Opportunistic Pathogenicity in an Invertebrate Model Host
- PMID: 29122853
- PMCID: PMC5765367
- DOI: 10.1534/g3.117.300245
Genome-Wide Screen for Saccharomyces cerevisiae Genes Contributing to Opportunistic Pathogenicity in an Invertebrate Model Host
Abstract
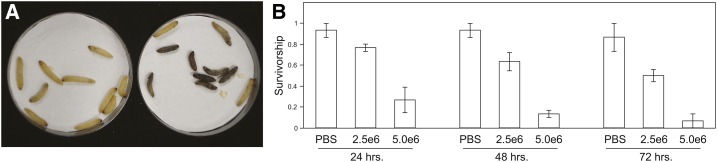
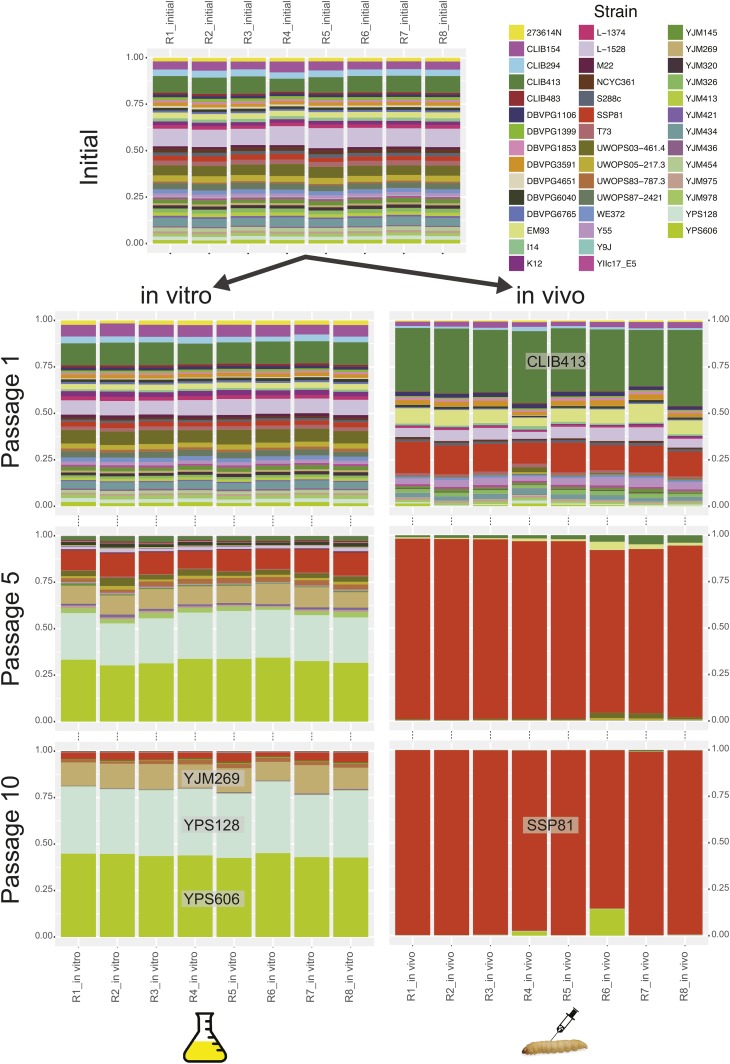
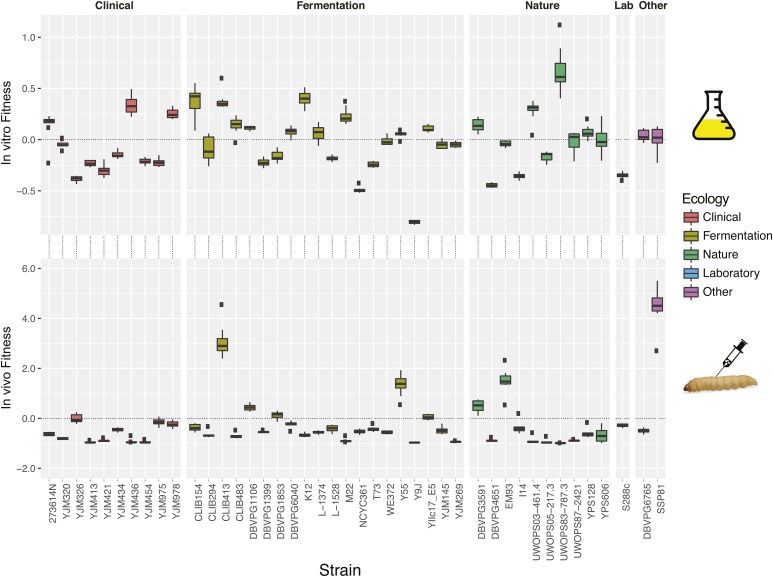
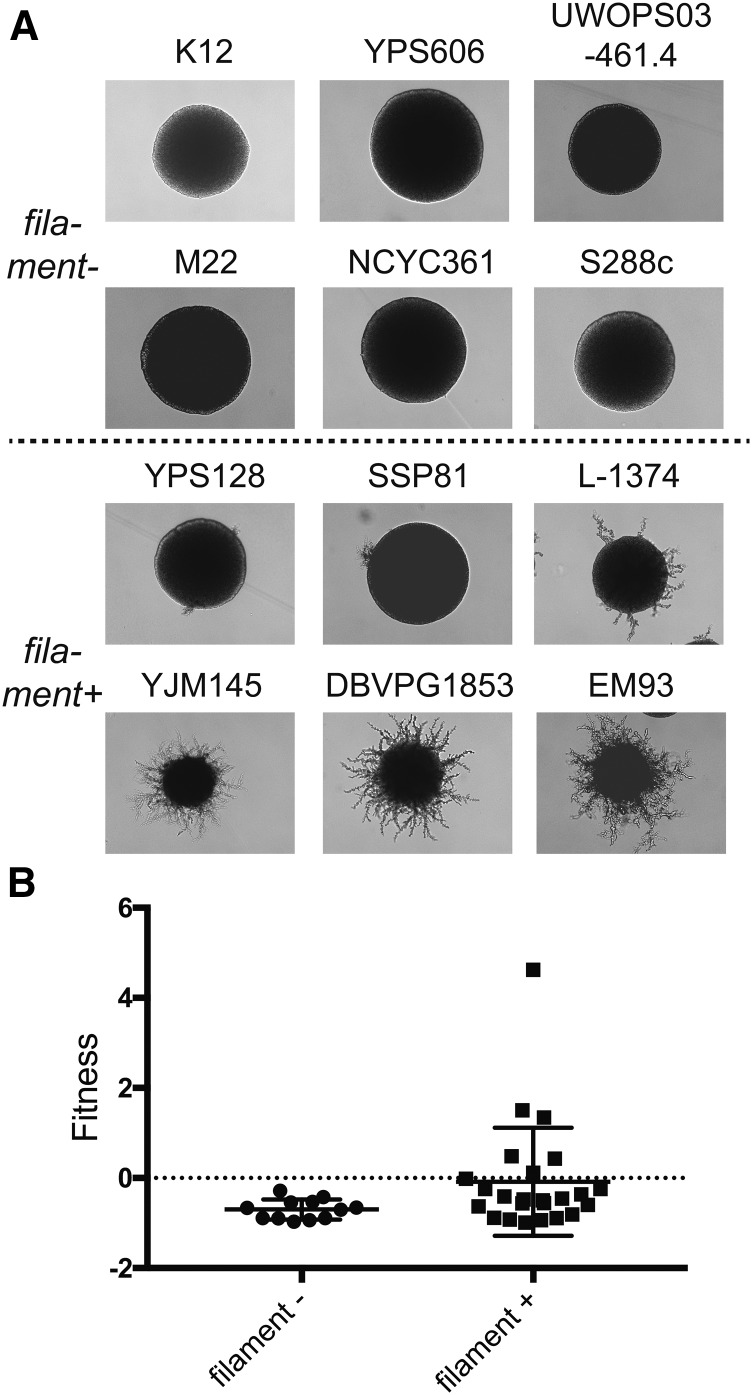
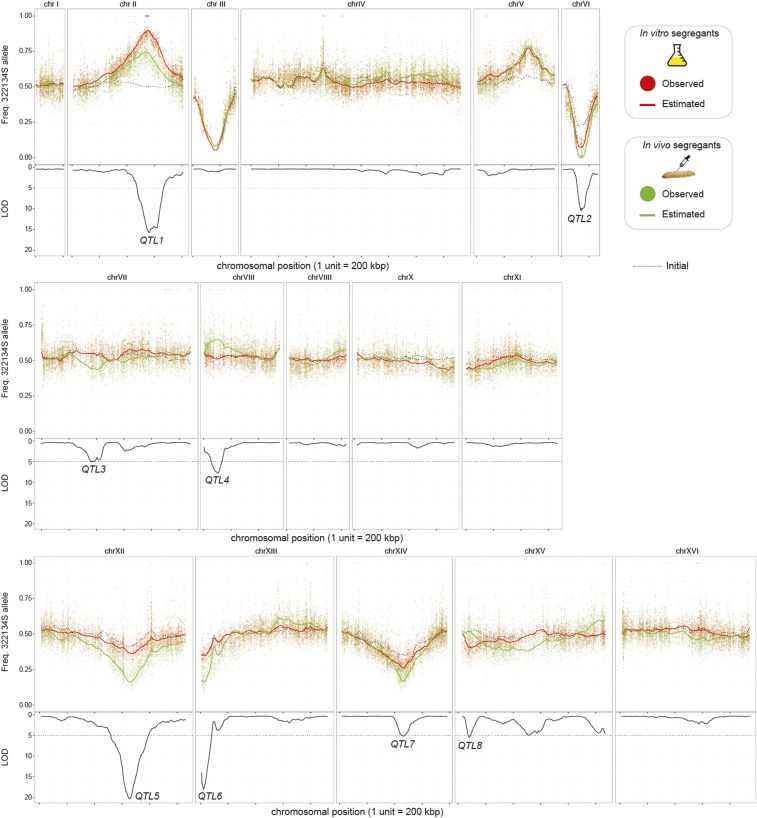
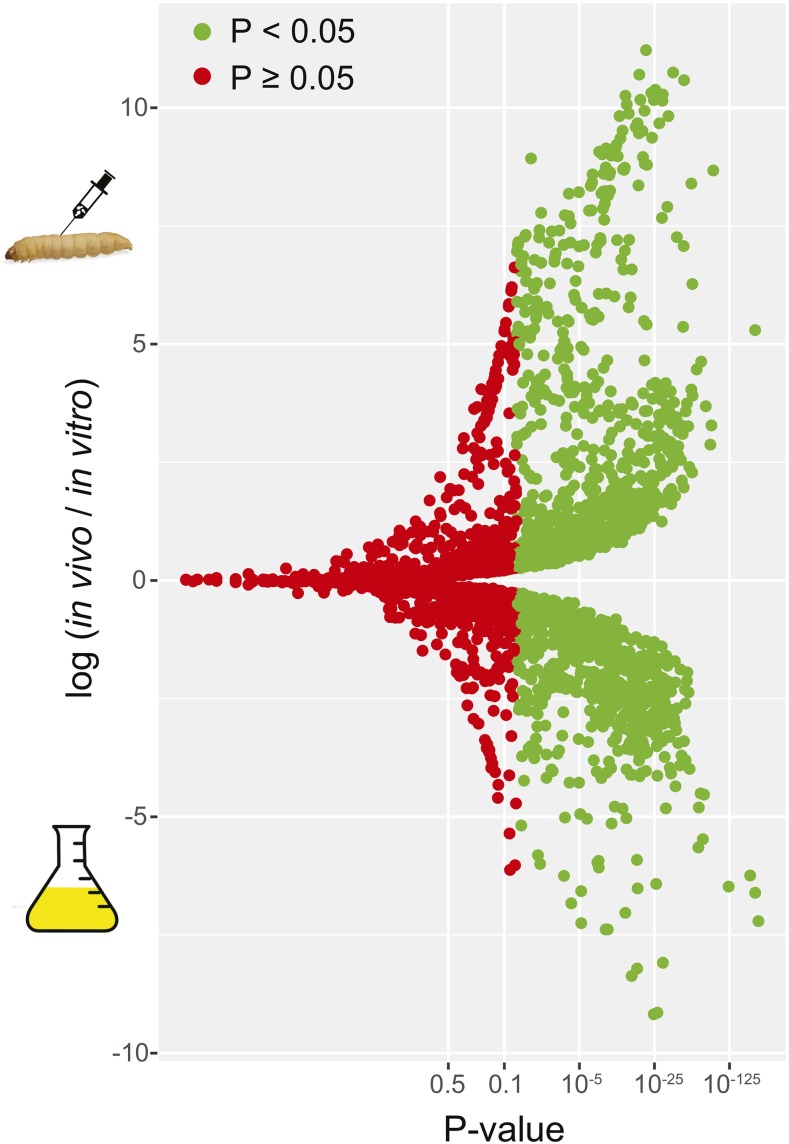
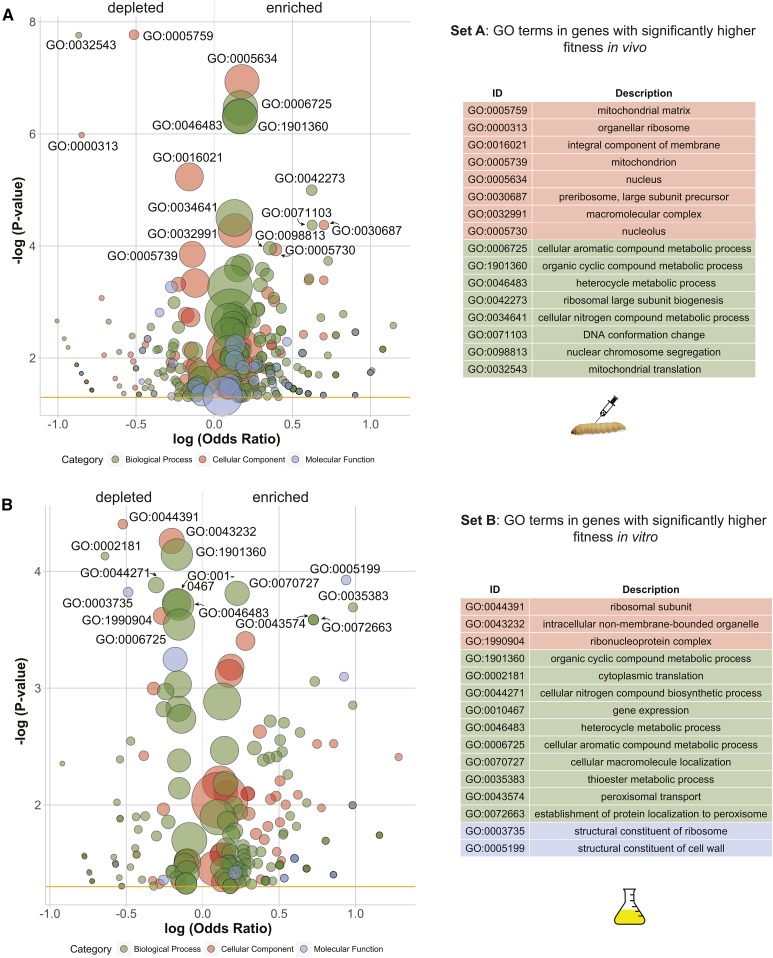
Environmental opportunistic pathogens can exploit vulnerable hosts through expression of traits selected for in their natural environments. Pathogenicity is itself a complicated trait underpinned by multiple complex traits, such as thermotolerance, morphology, and stress response. The baker's yeast, Saccharomyces cerevisiae, is a species with broad environmental tolerance that has been increasingly reported as an opportunistic pathogen of humans. Here we leveraged the genetic resources available in yeast and a model insect species, the greater waxmoth Galleria mellonella, to provide a genome-wide analysis of pathogenicity factors. Using serial passaging experiments of genetically marked wild-type strains, a hybrid strain was identified as the most fit genotype across all replicates. To dissect the genetic basis for pathogenicity in the hybrid isolate, bulk segregant analysis was performed which revealed eight quantitative trait loci significantly differing between the two bulks with alleles from both parents contributing to pathogenicity. A second passaging experiment with a library of deletion mutants for most yeast genes identified a large number of mutations whose relative fitness differed in vivovs.in vitro, including mutations in genes controlling cell wall integrity, mitochondrial function, and tyrosine metabolism. Yeast is presumably subjected to a massive assault by the innate insect immune system that leads to melanization of the host and to a large bottleneck in yeast population size. Our data support that resistance to the innate immune response of the insect is key to survival in the host and identifies shared genetic mechanisms between S. cerevisiae and other opportunistic fungal pathogens.
Keywords: barseq; bulk segregant analysis; hybrid; pseudohyphae; virulence.
Copyright © 2018 Phadke et al.
Figures
Similar articles
-
Oxidative stress survival in a clinical Saccharomyces cerevisiae isolate is influenced by a major quantitative trait nucleotide.Genetics. 2011 Jul;188(3):709-22. doi: 10.1534/genetics.111.128256. Epub 2011 Apr 21. Genetics. 2011. PMID: 21515583 Free PMC article.
-
Complex interactions of Klebsiella pneumoniae with the host immune system in a Galleria mellonella infection model.J Med Microbiol. 2013 Dec;62(Pt 12):1790-1798. doi: 10.1099/jmm.0.063032-0. Epub 2013 Sep 2. J Med Microbiol. 2013. PMID: 24000226
-
Conidiation color mutants of Aspergillus fumigatus are highly pathogenic to the heterologous insect host Galleria mellonella.PLoS One. 2009;4(1):e4224. doi: 10.1371/journal.pone.0004224. Epub 2009 Jan 19. PLoS One. 2009. PMID: 19156203 Free PMC article.
-
The greater wax moth Galleria mellonella: biology and use in immune studies.Pathog Dis. 2020 Nov 23;78(9):ftaa057. doi: 10.1093/femspd/ftaa057. Pathog Dis. 2020. PMID: 32970818 Free PMC article. Review.
-
Baker's Yeast Clinical Isolates Provide a Model for How Pathogenic Yeasts Adapt to Stress.Trends Genet. 2019 Nov;35(11):804-817. doi: 10.1016/j.tig.2019.08.002. Epub 2019 Sep 13. Trends Genet. 2019. PMID: 31526615 Free PMC article. Review.
Cited by
-
Impact of inter-species hybridisation on antifungal drug response in the Saccharomyces genus.BMC Genomics. 2024 Dec 2;25(1):1165. doi: 10.1186/s12864-024-11009-3. BMC Genomics. 2024. PMID: 39623390 Free PMC article.
-
Conserved signaling modules regulate filamentous growth in fungi: a model for eukaryotic cell differentiation.Genetics. 2024 Oct 7;228(2):iyae122. doi: 10.1093/genetics/iyae122. Genetics. 2024. PMID: 39239926 Review.
-
Standardization of G. mellonella Larvae to Provide Reliable and Reproducible Results in the Study of Fungal Pathogens.J Fungi (Basel). 2018 Sep 6;4(3):108. doi: 10.3390/jof4030108. J Fungi (Basel). 2018. PMID: 30200639 Free PMC article. Review.
-
Selection on plastic adherence leads to hyper-multicellular strains and incidental virulence in the budding yeast.Elife. 2023 Nov 2;12:e81056. doi: 10.7554/eLife.81056. Elife. 2023. PMID: 37916911 Free PMC article.
-
Complex genetics cause and constrain fungal persistence in different parts of the mammalian body.Genetics. 2022 Nov 1;222(3):iyac138. doi: 10.1093/genetics/iyac138. Genetics. 2022. PMID: 36103708 Free PMC article.
References
-
- Brennan M., Thomas D. Y., Whiteway M., Kavanagh K., 2002. Correlation between virulence of Candida albicans mutants in mice and Galleria mellonella larvae. FEMS Immunol. Med. Microbiol. 34: 153–157. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases