

SAMTOR is an S-adenosylmethionine sensor for the mTORC1 pathway
- PMID: 29123071
- PMCID: PMC5747364
- DOI: 10.1126/science.aao3265
SAMTOR is an S-adenosylmethionine sensor for the mTORC1 pathway
Abstract
mTOR complex 1 (mTORC1) regulates cell growth and metabolism in response to multiple environmental cues. Nutrients signal via the Rag guanosine triphosphatases (GTPases) to promote the localization of mTORC1 to the lysosomal surface, its site of activation. We identified SAMTOR, a previously uncharacterized protein, which inhibits mTORC1 signaling by interacting with GATOR1, the GTPase activating protein (GAP) for RagA/B. We found that the methyl donor S-adenosylmethionine (SAM) disrupts the SAMTOR-GATOR1 complex by binding directly to SAMTOR with a dissociation constant of approximately 7 μM. In cells, methionine starvation reduces SAM levels below this dissociation constant and promotes the association of SAMTOR with GATOR1, thereby inhibiting mTORC1 signaling in a SAMTOR-dependent fashion. Methionine-induced activation of mTORC1 requires the SAM binding capacity of SAMTOR. Thus, SAMTOR is a SAM sensor that links methionine and one-carbon metabolism to mTORC1 signaling.
Copyright © 2017 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works.
Figures
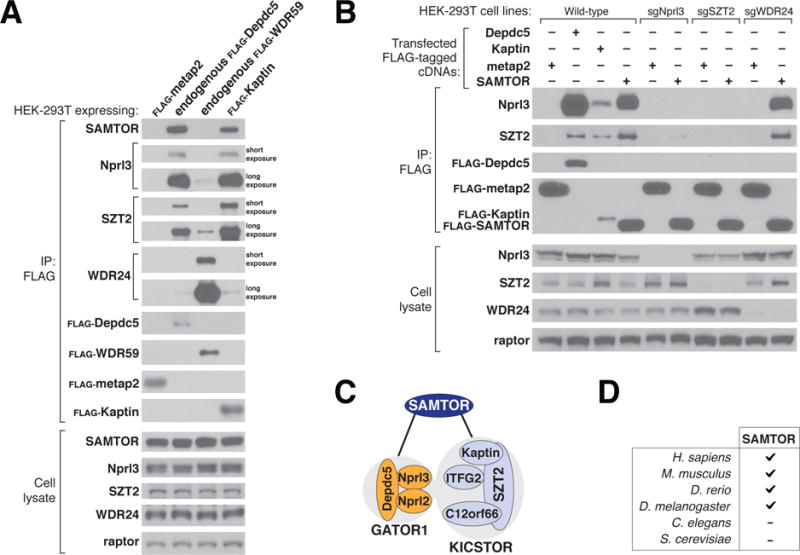
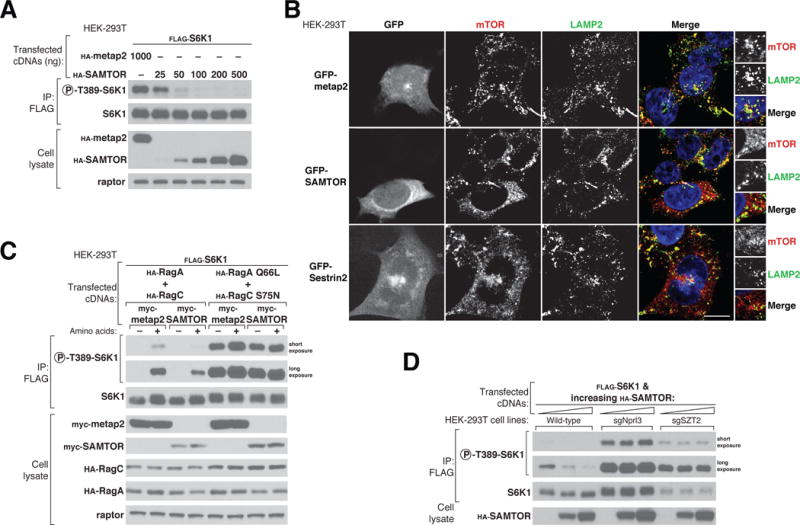
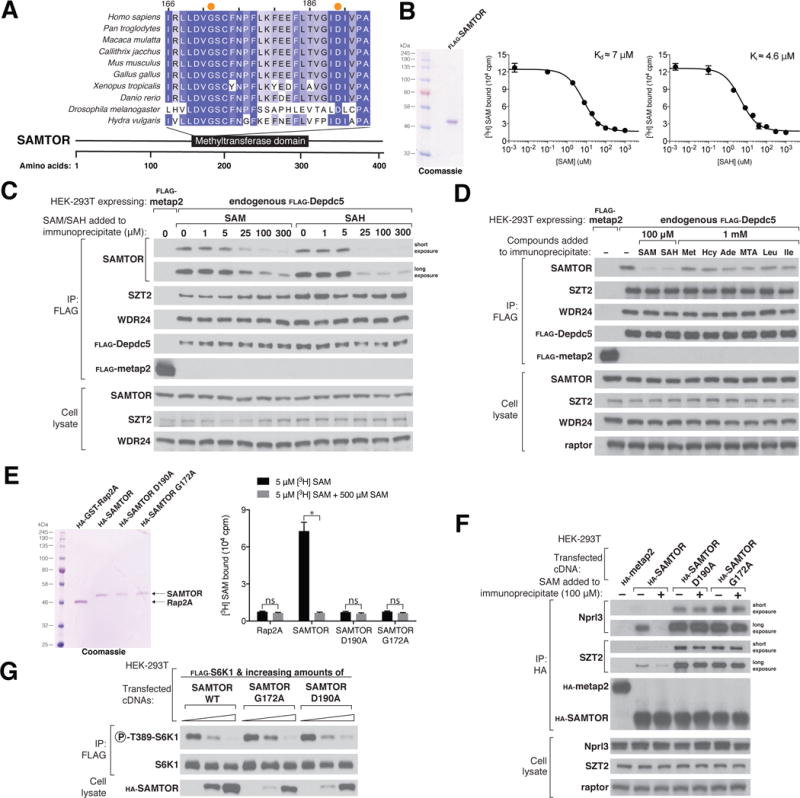
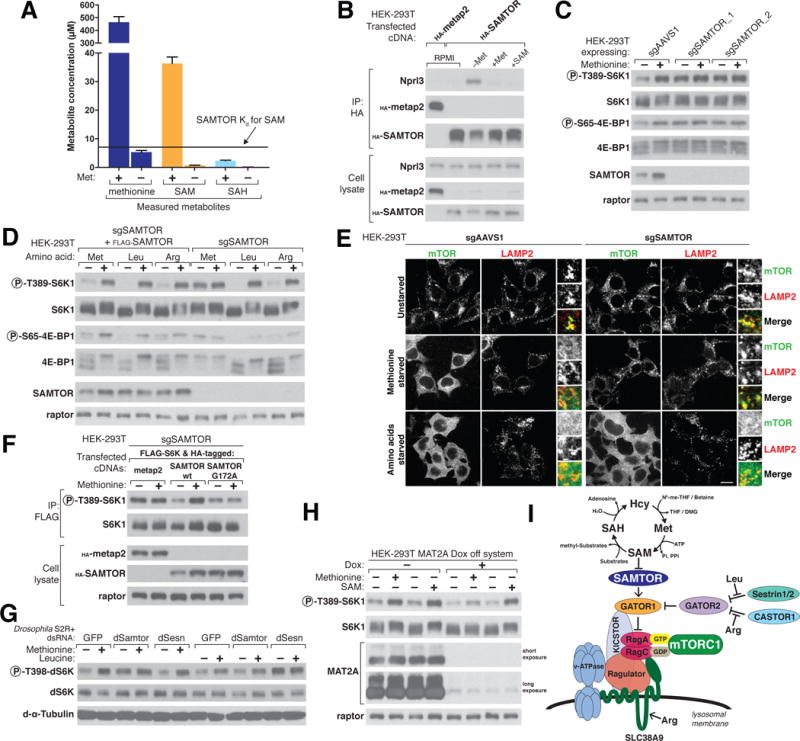
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
