

A Model-Based Approach for Separating the Cochlear Microphonic from the Auditory Nerve Neurophonic in the Ongoing Response Using Electrocochleography
- PMID: 29123468
- PMCID: PMC5662900
- DOI: 10.3389/fnins.2017.00592
A Model-Based Approach for Separating the Cochlear Microphonic from the Auditory Nerve Neurophonic in the Ongoing Response Using Electrocochleography
Abstract
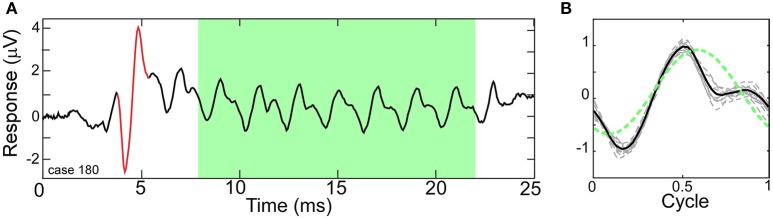
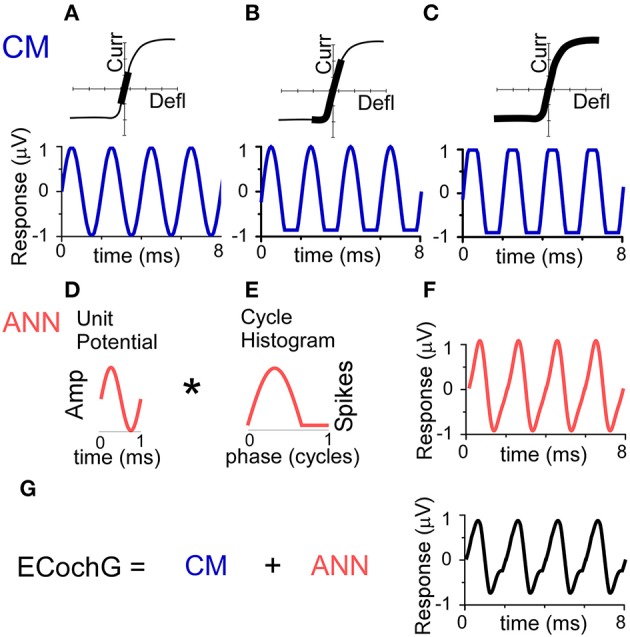
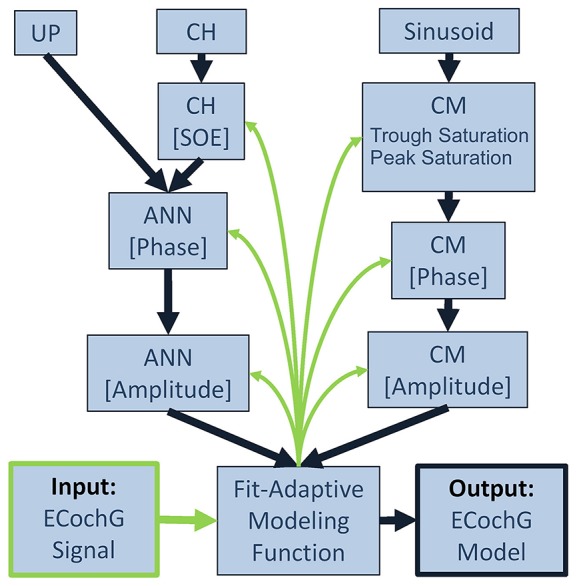
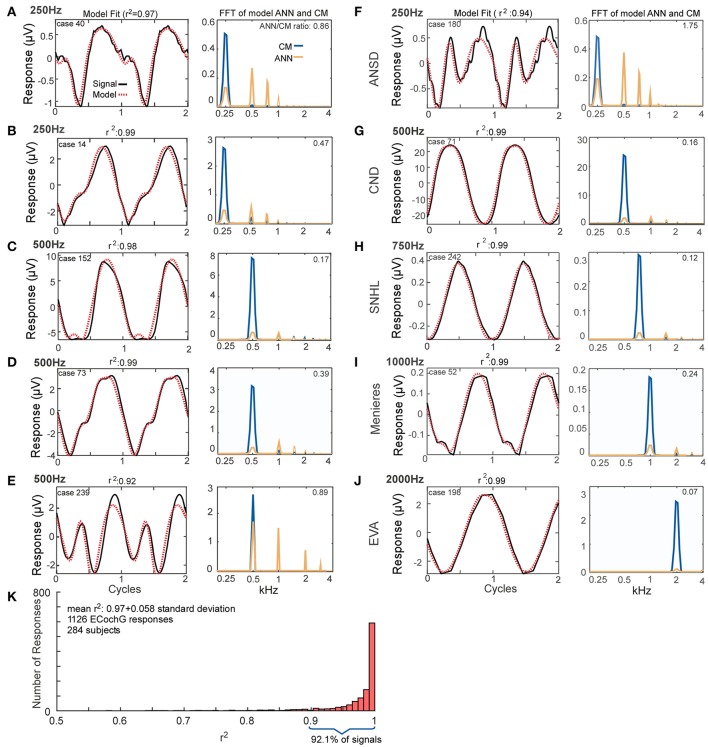
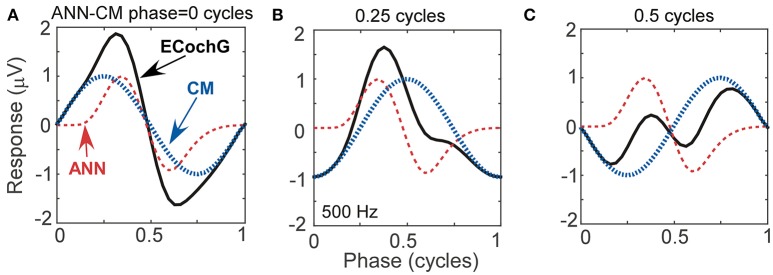
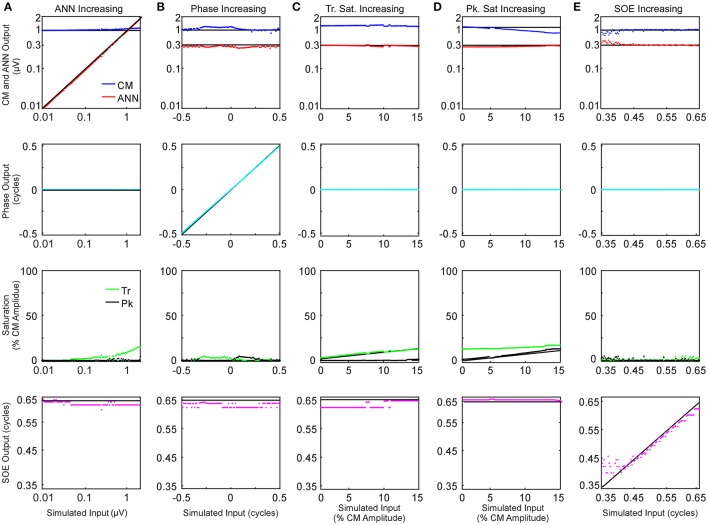
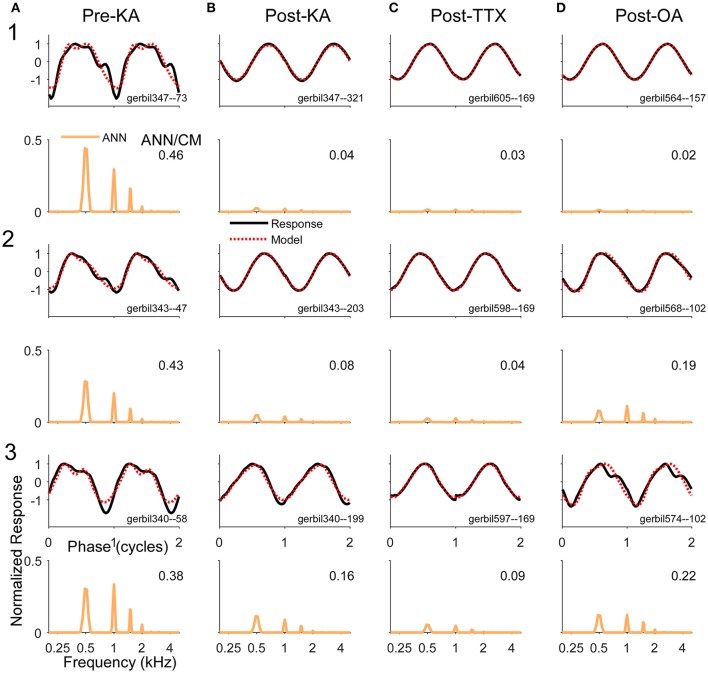
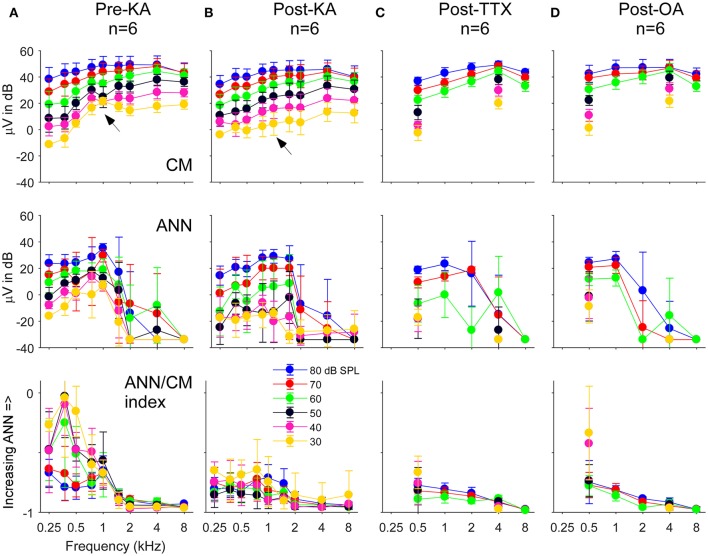
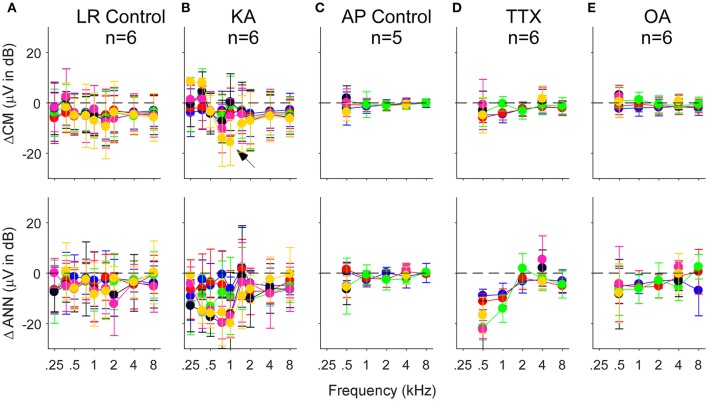
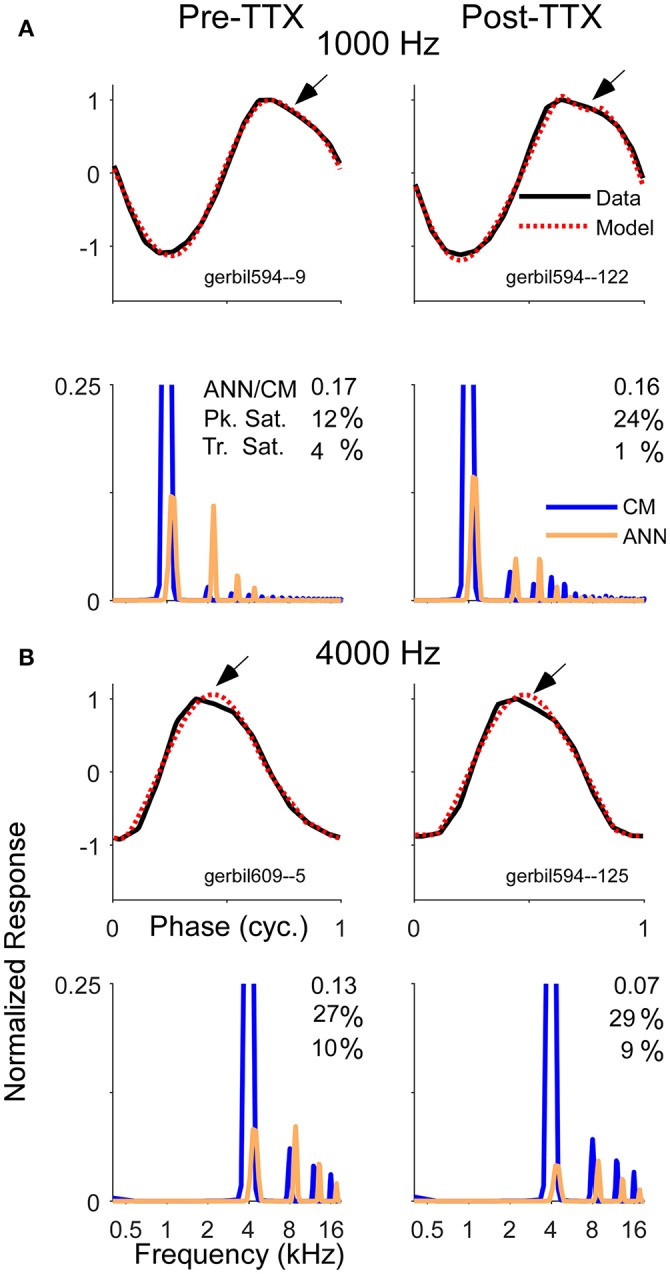
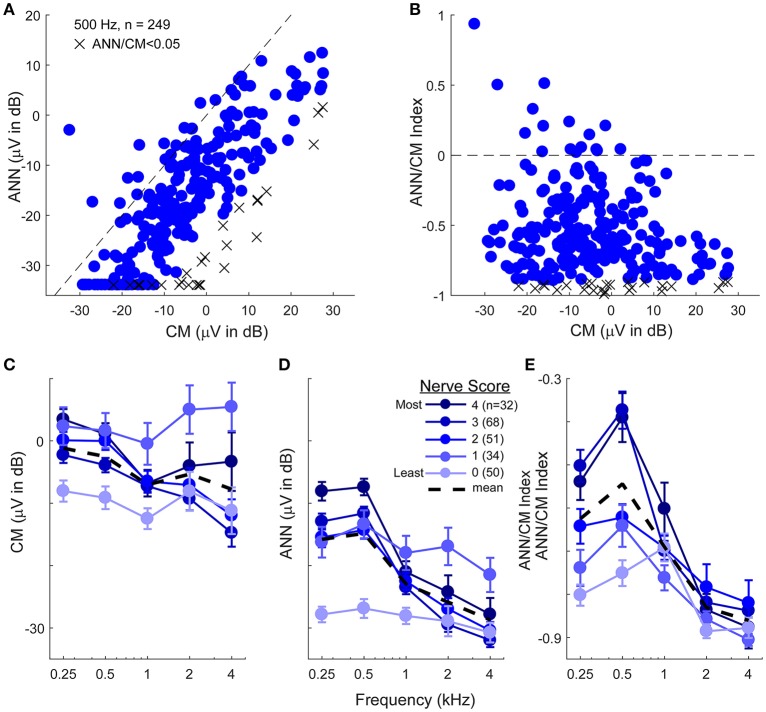
Electrocochleography (ECochG) is a potential clinically valuable technique for predicting speech perception outcomes in cochlear implant (CI) recipients, among other uses. Current analysis is limited by an inability to quantify hair cell and neural contributions which are mixed in the ongoing part of the response to low frequency tones. Here, we used a model based on source properties to account for recorded waveform shapes and to separate the combined signal into its components. The model for the cochlear microphonic (CM) was a sinusoid with parameters for independent saturation of the peaks and the troughs of the responses. The model for the auditory nerve neurophonic (ANN) was the convolution of a unit potential and population cycle histogram with a parameter for spread of excitation. Phases of the ANN and CM were additional parameters. The average cycle from the ongoing response was the input, and adaptive fitting identified CM and ANN parameters that best reproduced the waveform shape. Test datasets were responses recorded from the round windows of CI recipients, from the round window of gerbils before and after application of neurotoxins, and with simulated signals where each parameter could be manipulated in isolation. Waveforms recorded from 284 CI recipients had a variety of morphologies that the model fit with an average r2 of 0.97 ± 0.058 (standard deviation). With simulated signals, small systematic differences between outputs and inputs were seen with some variable combinations, but in general there were limited interactions among the parameters. In gerbils, the CM reported was relatively unaffected by the neurotoxins. In contrast, the ANN was strongly reduced and the reduction was limited to frequencies of 1,000 Hz and lower, consistent with the range of strong neural phase-locking. Across human CI subjects, the ANN contribution was variable, ranging from nearly none to larger than the CM. Development of this model could provide a means to isolate hair cell and neural activity that are mixed in the ongoing response to low-frequency tones. This tool can help characterize the residual physiology across CI subjects, and can be useful in other clinical settings where a description of the cochlear physiology is desirable.
Keywords: auditory hair cells; auditory nerve; auditory nerve model; cochlear physiology; computational modeling; electrophysiology; modeling and simulations.
Figures
Similar articles
-
Round Window Electrocochleography to Low Frequency Tones in Pediatric Cochlear Implant Recipients with and Without Auditory Neuropathy Spectrum Disorder: Separating Hair Cell and Neural Contributions Using a Computational Model.Otol Neurotol. 2022 Aug 1;43(7):781-788. doi: 10.1097/MAO.0000000000003568. Epub 2022 Jun 29. Otol Neurotol. 2022. PMID: 35763496 Free PMC article.
-
Postoperative Electrocochleography from Hybrid Cochlear Implant users: An Alternative Analysis Procedure.Hear Res. 2018 Dec;370:304-315. doi: 10.1016/j.heares.2018.10.016. Epub 2018 Oct 29. Hear Res. 2018. PMID: 30393003 Free PMC article.
-
Intraoperative Electrocochleographic Characteristics of Auditory Neuropathy Spectrum Disorder in Cochlear Implant Subjects.Front Neurosci. 2017 Jul 19;11:416. doi: 10.3389/fnins.2017.00416. eCollection 2017. Front Neurosci. 2017. PMID: 28769753 Free PMC article.
-
Audibility, speech perception and processing of temporal cues in ribbon synaptic disorders due to OTOF mutations.Hear Res. 2015 Dec;330(Pt B):200-12. doi: 10.1016/j.heares.2015.07.007. Epub 2015 Jul 15. Hear Res. 2015. PMID: 26188103 Review.
-
Cochlea and auditory nerve.Handb Clin Neurol. 2019;160:437-449. doi: 10.1016/B978-0-444-64032-1.00029-1. Handb Clin Neurol. 2019. PMID: 31277867 Review.
Cited by
-
Cochlear implants and other inner ear prostheses: today and tomorrow.Curr Opin Physiol. 2020 Dec;18:49-55. doi: 10.1016/j.cophys.2020.08.001. Epub 2020 Aug 14. Curr Opin Physiol. 2020. PMID: 32905432 Free PMC article.
-
Electrocochleography-Based Tonotopic Map: I. Place Coding of the Human Cochlea With Hearing Loss.Ear Hear. 2025 Jan-Feb 01;46(1):253-264. doi: 10.1097/AUD.0000000000001579. Epub 2024 Sep 5. Ear Hear. 2025. PMID: 39233326
-
Electrocochleography and cognition are important predictors of speech perception outcomes in noise for cochlear implant recipients.Sci Rep. 2022 Feb 23;12(1):3083. doi: 10.1038/s41598-022-07175-7. Sci Rep. 2022. PMID: 35197556 Free PMC article.
-
An analytic approach to identifying the sources of the low-frequency round window cochlear response.Hear Res. 2019 Apr;375:53-65. doi: 10.1016/j.heares.2019.02.001. Epub 2019 Feb 15. Hear Res. 2019. PMID: 30808536 Free PMC article.
-
Middle ear muscle and medial olivocochlear activity inferred from individual human ears via cochlear potentials.J Acoust Soc Am. 2023 Mar;153(3):1723. doi: 10.1121/10.0017604. J Acoust Soc Am. 2023. PMID: 37002081 Free PMC article.
References
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources