

Taurine as an Essential Neuromodulator during Perinatal Cortical Development
- PMID: 29123472
- PMCID: PMC5662885
- DOI: 10.3389/fncel.2017.00328
Taurine as an Essential Neuromodulator during Perinatal Cortical Development
Abstract
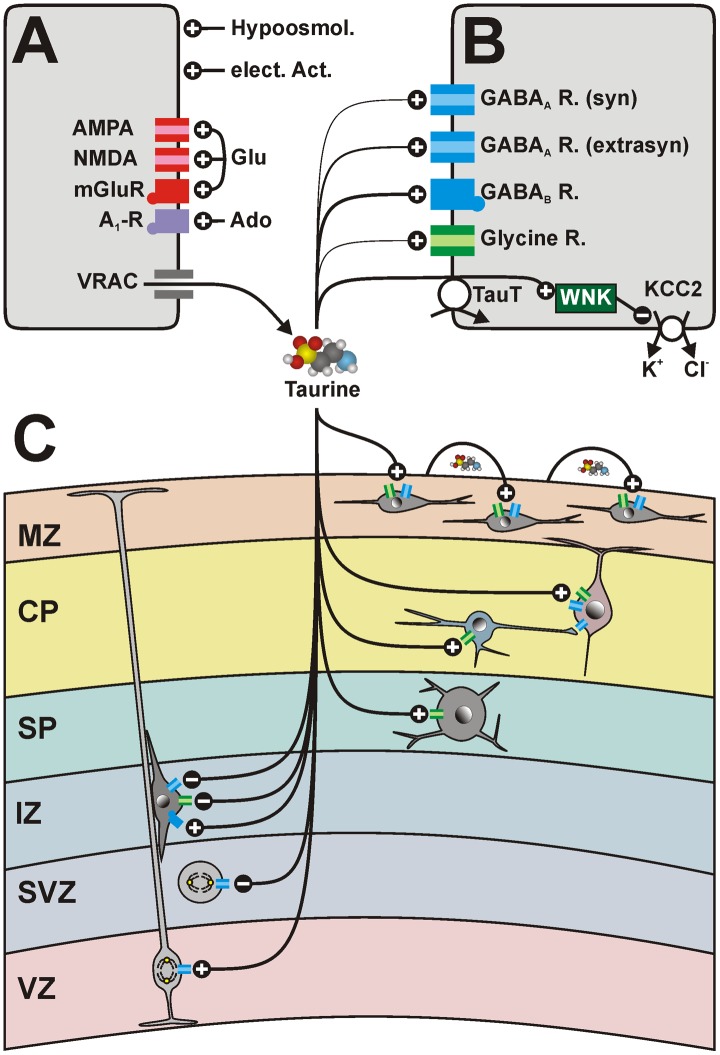
A variety of experimental studies demonstrated that neurotransmitters are an important factor for the development of the central nervous system, affecting neurodevelopmental events like neurogenesis, neuronal migration, programmed cell death, and differentiation. While the role of the classical neurotransmitters glutamate and gamma-aminobutyric acid (GABA) on neuronal development is well established, the aminosulfonic acid taurine has also been considered as possible neuromodulator during early neuronal development. The purpose of the present review article is to summarize the properties of taurine as neuromodulator in detail, focusing on the direct involvement of taurine on various neurodevelopmental events and the regulation of neuronal activity during early developmental epochs. The current knowledge is that taurine lacks a synaptic release mechanism but is released by volume-sensitive organic anion channels and/or a reversal of the taurine transporter. Extracellular taurine affects neurons and neuronal progenitor cells mainly via glycine, GABA(A), and GABA(B) receptors with considerable receptor and subtype-specific affinities. Taurine has been shown to directly influence neurogenesis in vitro as well as neuronal migration in vitro and in vivo. It provides a depolarizing signal for a variety of neuronal population in the immature central nervous system, thereby directly influencing neuronal activity. While in the neocortex, taurine probably enhance neuronal activity, in the immature hippocampus, a tonic taurinergic tone might be necessary to attenuate activity. In summary, taurine must be considered as an essential modulator of neurodevelopmental events, and possible adverse consequences on fetal and/or early postnatal development should be evaluated for pharmacological therapies affecting taurinergic functions.
Keywords: Cajal–Retzius cells; GABA receptors; cerebral cortex; glycine receptors; migration; review; rodent; subplate.
Figures
Similar articles
-
Putative Role of Taurine as Neurotransmitter During Perinatal Cortical Development.Adv Exp Med Biol. 2017;975 Pt 1:281-292. doi: 10.1007/978-94-024-1079-2_25. Adv Exp Med Biol. 2017. PMID: 28849463
-
Maternal taurine as a modulator of Cl- homeostasis as well as of glycine/GABAA receptors for neocortical development.Front Cell Neurosci. 2023 Aug 3;17:1221441. doi: 10.3389/fncel.2023.1221441. eCollection 2023. Front Cell Neurosci. 2023. PMID: 37601283 Free PMC article. Review.
-
Prenatal and postnatal contents of amino acid neurotransmitters in mouse parietal cortex.Dev Neurosci. 2003 Sep-Oct;25(5):366-74. doi: 10.1159/000073514. Dev Neurosci. 2003. PMID: 14614264
-
Depolarizing glycine responses in Cajal-Retzius cells of neonatal rat cerebral cortex.Neuroscience. 2002;112(2):299-307. doi: 10.1016/s0306-4522(02)00071-4. Neuroscience. 2002. PMID: 12044448
-
Spontaneous Neuronal Activity in Developing Neocortical Networks: From Single Cells to Large-Scale Interactions.Front Neural Circuits. 2016 May 24;10:40. doi: 10.3389/fncir.2016.00040. eCollection 2016. Front Neural Circuits. 2016. PMID: 27252626 Free PMC article. Review.
Cited by
-
Taurine depletion during fetal and postnatal development blunts firing responses of neocortical layer II/III pyramidal neurons.Front Mol Neurosci. 2022 Nov 17;15:806798. doi: 10.3389/fnmol.2022.806798. eCollection 2022. Front Mol Neurosci. 2022. PMID: 36466806 Free PMC article.
-
Modulation of Neocortical Development by Early Neuronal Activity: Physiology and Pathophysiology.Front Cell Neurosci. 2017 Nov 29;11:379. doi: 10.3389/fncel.2017.00379. eCollection 2017. Front Cell Neurosci. 2017. PMID: 29238291 Free PMC article. Review.
-
The effects of electrical stimulation of the peripheral vestibular system on neurochemical release in the rat striatum.PLoS One. 2018 Oct 29;13(10):e0205869. doi: 10.1371/journal.pone.0205869. eCollection 2018. PLoS One. 2018. PMID: 30372453 Free PMC article.
-
Taurine and Astrocytes: A Homeostatic and Neuroprotective Relationship.Front Mol Neurosci. 2022 Jul 5;15:937789. doi: 10.3389/fnmol.2022.937789. eCollection 2022. Front Mol Neurosci. 2022. PMID: 35866158 Free PMC article. Review.
-
Longitudinal Biochemical and Behavioral Alterations in a Gyrencephalic Model of Blast-Related Mild Traumatic Brain Injury.Neurotrauma Rep. 2024 Mar 14;5(1):254-266. doi: 10.1089/neur.2024.0002. eCollection 2024. Neurotrauma Rep. 2024. PMID: 38515547 Free PMC article.
References
-
- Ahring P. K., Bang L. H., Jensen M. L., Strobaek D., Hartiadi L. Y., Chebib M., et al. (2016). A pharmacological assessment of agonists and modulators at α4β2γ2 and α4β2δ GABAA receptors: The challenge in comparing apples with oranges. Pharmacol. Res. 111 563–576. 10.1016/j.phrs.2016.05.014 - DOI - PubMed
-
- Altshuler D., LoTurco J. J., Rush J., Cepko C. (1993). Taurine promotes the differentiation of a vertebrate retinal cell type in vitro. Development 119 1317–1328. - PubMed
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources