

The telomere bouquet facilitates meiotic prophase progression and exit in fission yeast
- PMID: 29123917
- PMCID: PMC5674143
- DOI: 10.1038/celldisc.2017.41
The telomere bouquet facilitates meiotic prophase progression and exit in fission yeast
Abstract
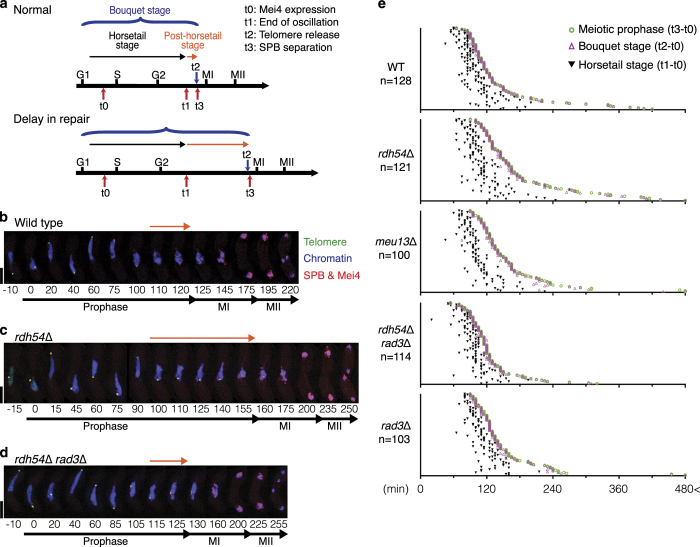
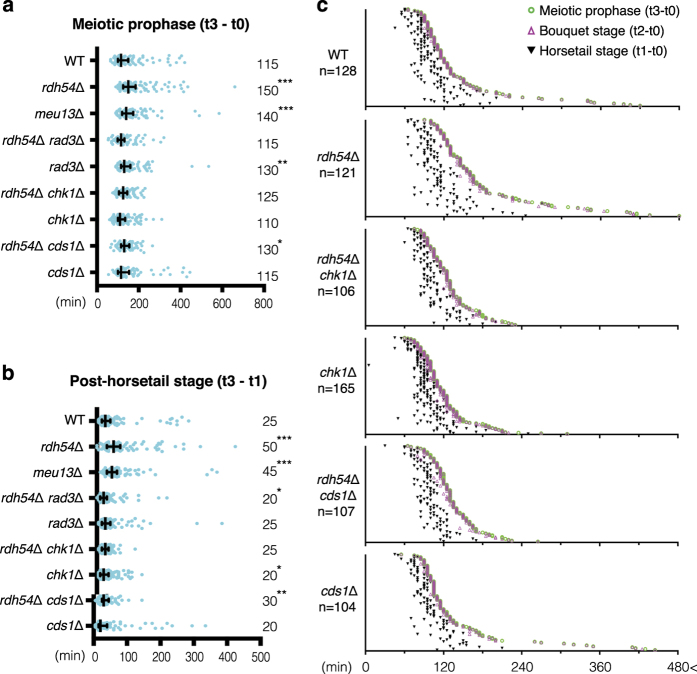
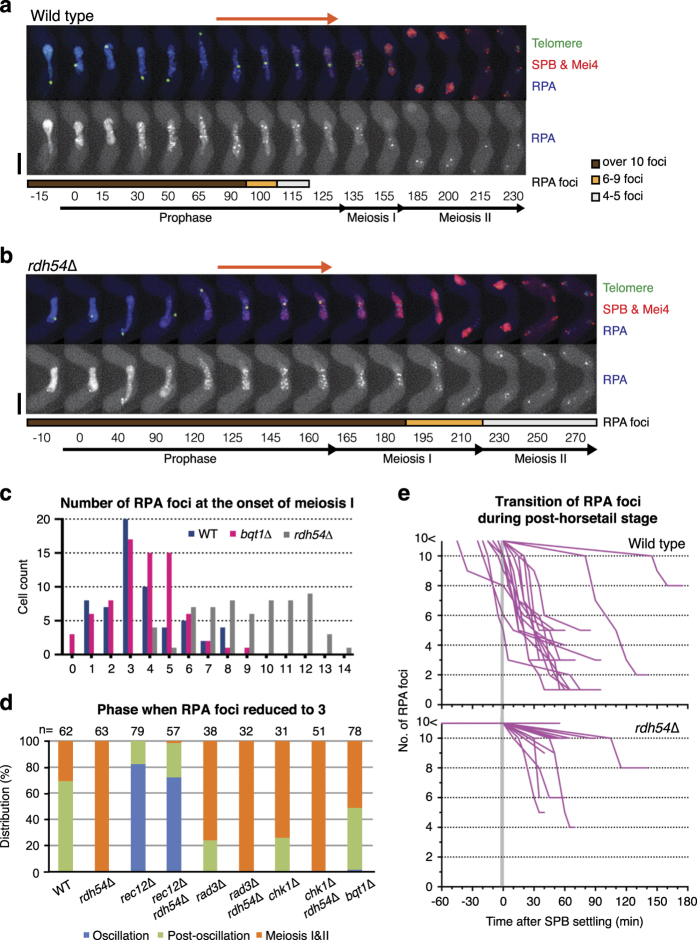
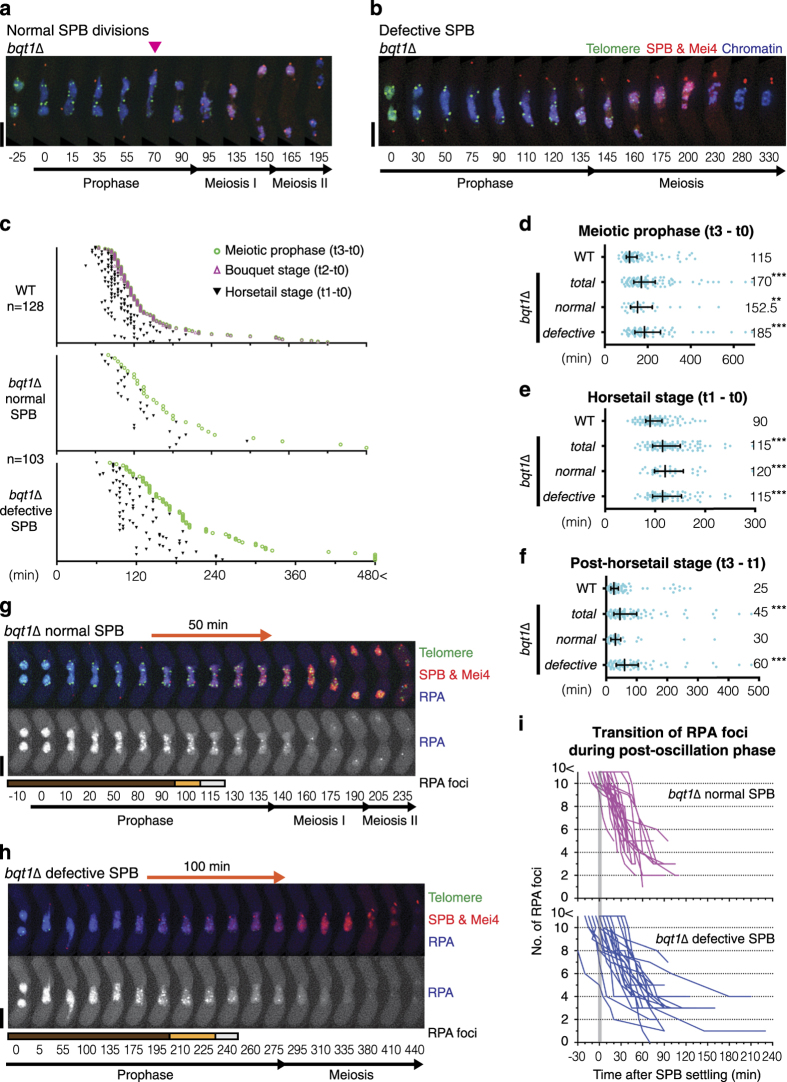
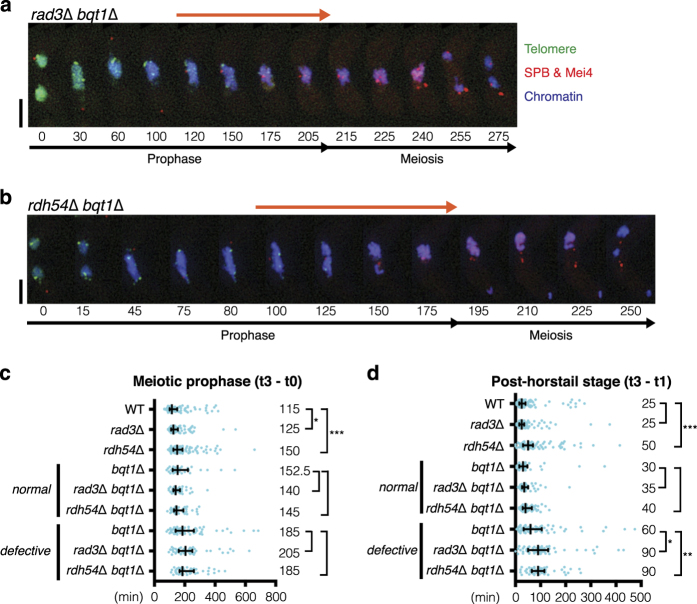
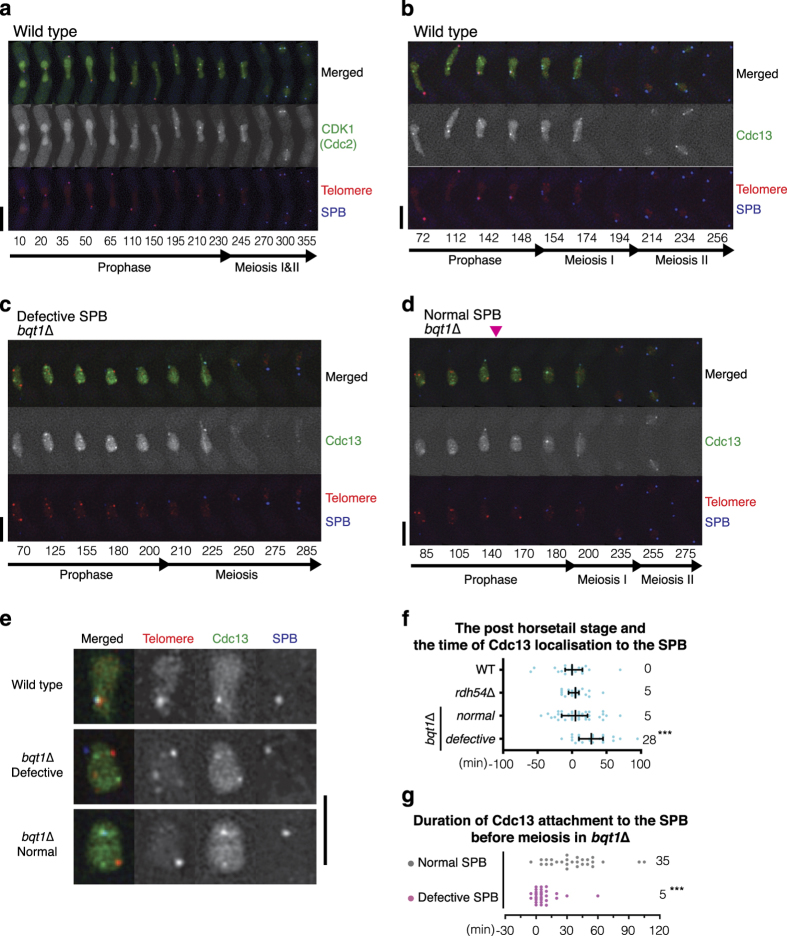
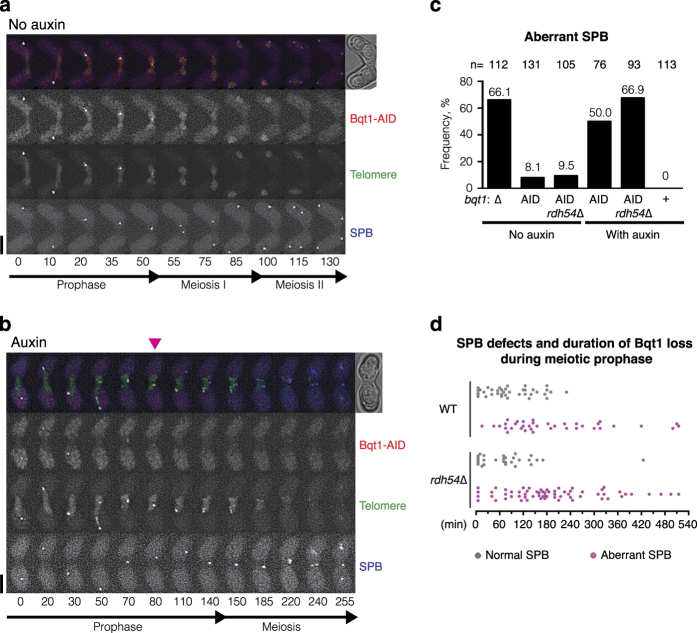
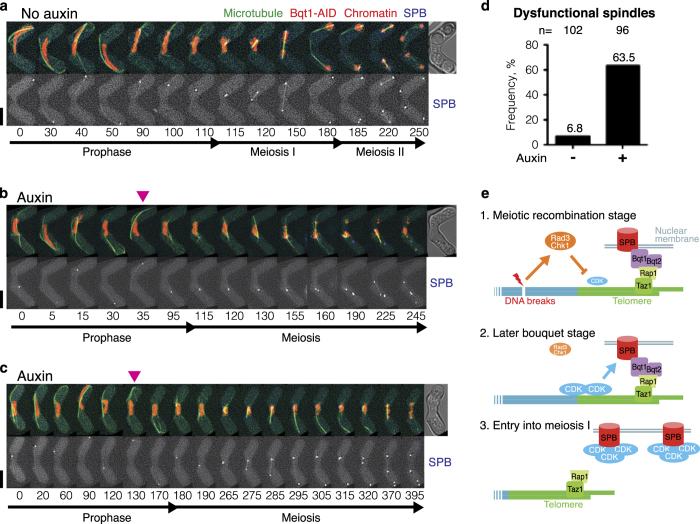
During meiotic prophase, chromosome arrangement and oscillation promote the pairing of homologous chromosomes for meiotic recombination. This dramatic movement involves clustering of telomeres at the nuclear membrane to form the so-called telomere bouquet. In fission yeast, the telomere bouquet is formed near the spindle pole body (SPB), which is the microtubule organising centre, functionally equivalent to the metazoan centrosome. Disruption of bouquet configuration impedes homologous chromosome pairing, meiotic recombination and spindle formation. Here, we demonstrate that the bouquet is maintained throughout meiotic prophase and promotes timely prophase exit in fission yeast. Persistent DNA damages, induced during meiotic recombination, activate the Rad3 and Chk1 DNA damage checkpoint kinases and extend the bouquet stage beyond the chromosome oscillation period. The auxin-inducible degron system demonstrated that premature termination of the bouquet stage leads to severe extension of prophase and consequently spindle formation defects. However, this delayed exit from meiotic prophase was not caused by residual DNA damage. Rather, loss of chromosome contact with the SPB caused delayed accumulation of CDK1-cyclin B at the SPB, which correlated with impaired SPB separation. In the absence of the bouquet, CDK1-cyclin B localised near the telomeres but not at the SPB at the later stage of meiotic prophase. Thus, bouquet configuration is maintained throughout meiotic prophase, by which this spatial organisation may facilitate local and timely activation of CDK1 near the SPB. Our findings illustrate that chromosome contact with the nuclear membrane synchronises meiotic progression of the nucleoplasmic chromosomes with that of the cytoplasmic SPB.
Keywords: Cdc2-Cdc13; Rad3–Chk1; S. pombe; bouquet; chromosome; meiosis; telomeres; the LINC complex.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
A Taz1- and Microtubule-Dependent Regulatory Relationship between Telomere and Centromere Positions in Bouquet Formation Secures Proper Meiotic Divisions.PLoS Genet. 2016 Sep 9;12(9):e1006304. doi: 10.1371/journal.pgen.1006304. eCollection 2016 Sep. PLoS Genet. 2016. PMID: 27611693 Free PMC article.
-
Meiotic proteins bqt1 and bqt2 tether telomeres to form the bouquet arrangement of chromosomes.Cell. 2006 Apr 7;125(1):59-69. doi: 10.1016/j.cell.2006.01.048. Cell. 2006. PMID: 16615890
-
Bqt2p is essential for initiating telomere clustering upon pheromone sensing in fission yeast.J Cell Biol. 2006 Jun 19;173(6):845-51. doi: 10.1083/jcb.200602152. Epub 2006 Jun 12. J Cell Biol. 2006. PMID: 16769823 Free PMC article.
-
A bouquet of chromosomes.J Cell Sci. 2004 Aug 15;117(Pt 18):4025-32. doi: 10.1242/jcs.01363. J Cell Sci. 2004. PMID: 15316078 Review.
-
Homologous chromosome pairing in Schizosaccharomyces pombe.Yeast. 2006 Oct 15;23(13):977-89. doi: 10.1002/yea.1403. Yeast. 2006. PMID: 17072890 Review.
Cited by
-
Rewiring for movements in meiotic prophase: regulators, roles, and evolutionary pathways.Curr Opin Genet Dev. 2025 Aug;93:102366. doi: 10.1016/j.gde.2025.102366. Epub 2025 Jun 7. Curr Opin Genet Dev. 2025. PMID: 40484002 Review.
-
Formation of the synaptonemal complex in a gynogenetic allodiploid hybrid fish.Front Genet. 2023 Feb 27;14:998775. doi: 10.3389/fgene.2023.998775. eCollection 2023. Front Genet. 2023. PMID: 36923790 Free PMC article.
-
A visual atlas of meiotic protein dynamics in living fission yeast.Open Biol. 2021 Feb;11(2):200357. doi: 10.1098/rsob.200357. Epub 2021 Feb 24. Open Biol. 2021. PMID: 33622106 Free PMC article.
-
Identifying Chromosome Movement Patterns During Meiosis Using ChroMo.Methods Mol Biol. 2024;2818:271-288. doi: 10.1007/978-1-0716-3906-1_18. Methods Mol Biol. 2024. PMID: 39126481
-
Cyclins and CDKs in the regulation of meiosis-specific events.Front Cell Dev Biol. 2022 Nov 29;10:1069064. doi: 10.3389/fcell.2022.1069064. eCollection 2022. Front Cell Dev Biol. 2022. PMID: 36523509 Free PMC article. Review.
References
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
