

Influence of chemical denaturants on the activity, fold and zinc status of anthrax lethal factor
- PMID: 29124135
- PMCID: PMC5668564
- DOI: 10.1016/j.bbrep.2015.03.004
Influence of chemical denaturants on the activity, fold and zinc status of anthrax lethal factor
Abstract
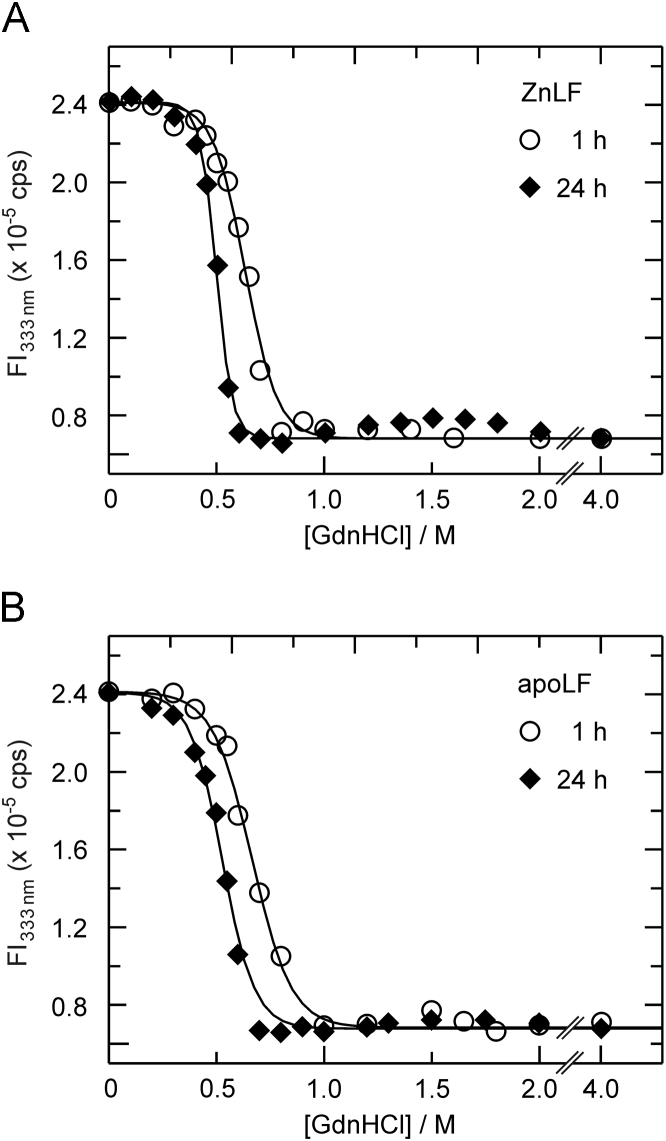
Anthrax lethal factor (LF) is a zinc-dependent endopeptidase which, through a process facilitated by protective antigen, translocates to the host cell cytosol in a partially unfolded state. In the current report, the influence of urea and guanidine hydrochloride (GdnHCl) on LF׳s catalytic function, fold and metal binding was assessed at neutral pH. Both urea and GdnHCl were found to inhibit LF prior to the onset of unfolding, with the inhibition by the latter denaturant being a consequence of its ionic strength. With the exception of demetallated LF (apoLF) in urea, unfolding, as monitored by tryptophan fluorescence spectroscopy, was found to follow a two-state (native to unfolded) mechanism. Analysis of the metal status of LF with 4-(2-pyridylazoresorcinol) (PAR) following urea or GdnHCl exposure suggests the enzyme to be capable of maintaining its metal ion passed the observed unfolding transition in a chelator-inaccessible form. Although an increase in the concentration of the denaturants eventually allowed the chelator access to the protein׳s zinc ion, such process is not correlated with the release of the metal ion. Indeed, significant dissociation of the zinc ion from LF was not observed even at 6 M urea, and only high concentrations of GdnHCl (>3 M) were capable of inducing the release of the metal ion from the protein. Hence, the current study demonstrates not only the propensity of LF to tightly bind its zinc ion beyond the spectroscopically determined unfolding transition, but also the utility of PAR as a structural probe.
Keywords: 4-(2-pyridylazo)resorcinol; CD, circular dichroism; Chemical denaturants; DPA, dipicolinic acid; EDTA, ethylenediaminetetraacetic acid; EF, edema factor; LF, anthrax lethal factor; Lethal factor; MWCO, molecular weight cut-off; PA, protective antigen; PAR, 4-(2-pyridylazo)resorcinol; Protein folding; S-pNA, lethal factor substrate; SASA, solvent-accessible surface area; SOD, superoxide dismutase; Tryptophan fluorescence; Zinc; ZnLF, zinc-containing lethal factor; cps, counts per second.
Figures





References
-
- Mock M., Fouet A. Anthrax. Annu. Rev. Microbiol. 2001;55:647–671. - PubMed
-
- Collier R.J., Young J.A. Anthrax toxin. Annu. Rev. Cell Dev. Biol. 2003;19:45–70. - PubMed
-
- Young J.A., Collier R.J. Anthrax toxin: receptor binding, internalization, pore formation, and translocation. Annu. Rev. Biochem. 2007;76:243–265. - PubMed
-
- Petosa C., Collier R.J., Klimpel K.R., Leppla S.H., Liddington R.C. Crystal structure of the anthrax toxin protective antigen. Nature. 1997;385:833–838. - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
