

Enhancing hypothiocyanite production by lactoperoxidase - mechanism and chemical properties of promotors
- PMID: 29124212
- PMCID: PMC5669353
- DOI: 10.1016/j.bbrep.2015.10.001
Enhancing hypothiocyanite production by lactoperoxidase - mechanism and chemical properties of promotors
Abstract
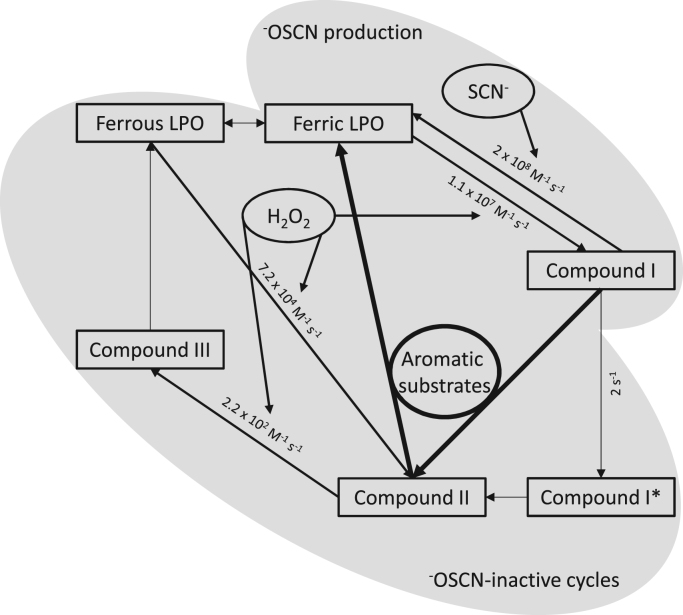
Background: The heme enzyme lactoperoxidase is found in body secretions where it significantly contributes to the humoral immune response against pathogens. After activation the peroxidase oxidizes thiocyanate to hypothiocyanite which is known for its microbicidal properties. Yet several pathologies are accompanied by a disturbed hypothiocyanite production which results in a reduced immune defense.
Methods: The results were obtained by measuring enzyme-kinetic parameters using UV-vis spectroscopy and a standardized enzyme-kinetic test system as well as by the determination of second order rate constants using stopped-flow spectroscopy.
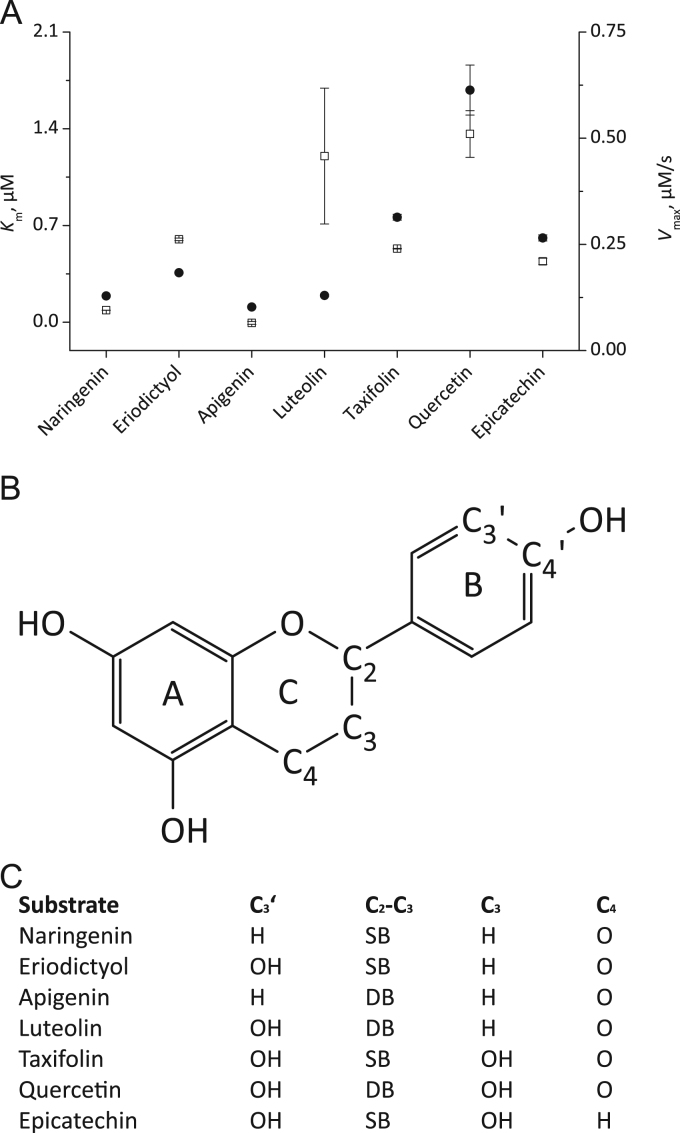
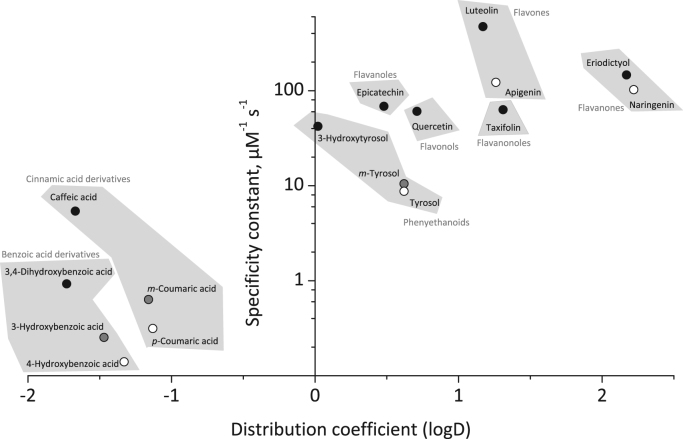
Results: In this study we systematically tested thirty aromatic substrates for their efficiency to promote the lactoperoxidase-mediated hypothiocyanite production by restoring the native ferric enzyme state. Thereby hydrophobic compounds with a 3,4-dihydroxyphenyl partial structure such as hydroxytyrosol and selected flavonoids emerged as highly efficient promotors of the (pseudo-)halogenating lactoperoxidase activity.
Conclusions: This study discusses important structure-function relationships of efficient aromatic LPO substrates and may contribute to the development of new agents to promote lactoperoxidase activity in secretory fluids of patients.
Significance: This study may contribute to a better understanding of the (patho-)physiological importance of the (pseudo-)halogenating lactoperoxidase activity. The presented results may in future lead to the development of new therapeutic strategies which, by reactivating lactoperoxidase-derived hypothiocyanite production, promote the immunological activity of this enzyme.
Keywords: 3,4-dihydroxylated compounds; ABTS, 2,2′-azinobis(3-ethylbenzothiazoline-6-sulfonic acid); Aromatic compounds; DB, double bond; DTNB, 5,5′-dithiobis-(2-nitrobenzoic acid); EPO, eosinophil peroxidase; Hypothiocyanite; Inflammation; LPO, lactoperoxidase; Lactoperoxidase; MPO, myeloperoxidase; Peroxidases; SB, single bond; TNB, 5-thio-2-nitrobenzoic acid.; ssp., subspecies.
Figures




Similar articles
-
Flavonoids as promoters of the (pseudo-)halogenating activity of lactoperoxidase and myeloperoxidase.Free Radic Biol Med. 2016 Aug;97:307-319. doi: 10.1016/j.freeradbiomed.2016.06.026. Epub 2016 Jun 25. Free Radic Biol Med. 2016. PMID: 27350402
-
Lactoperoxidase as a potential drug target.Expert Opin Ther Targets. 2016;20(4):447-61. doi: 10.1517/14728222.2016.1112378. Epub 2015 Nov 11. Expert Opin Ther Targets. 2016. PMID: 26558497 Review.
-
Components of a standardised olive leaf dry extract (Ph. Eur.) promote hypothiocyanite production by lactoperoxidase.Arch Biochem Biophys. 2014 May 1;549:17-25. doi: 10.1016/j.abb.2014.03.006. Epub 2014 Mar 18. Arch Biochem Biophys. 2014. PMID: 24657078
-
Uric acid and thiocyanate as competing substrates of lactoperoxidase.J Biol Chem. 2014 Aug 8;289(32):21937-49. doi: 10.1074/jbc.M113.544957. Epub 2014 Jun 13. J Biol Chem. 2014. PMID: 24928513 Free PMC article.
-
Heme Peroxidases at Unperturbed and Inflamed Mucous Surfaces.Antioxidants (Basel). 2021 Nov 12;10(11):1805. doi: 10.3390/antiox10111805. Antioxidants (Basel). 2021. PMID: 34829676 Free PMC article. Review.
Cited by
-
Inorganic Salts and Antimicrobial Photodynamic Therapy: Mechanistic Conundrums?Molecules. 2018 Dec 3;23(12):3190. doi: 10.3390/molecules23123190. Molecules. 2018. PMID: 30514001 Free PMC article. Review.
-
Antibacterial and antiplaque efficacy of a lactoperoxidase-thiocyanate-hydrogen-peroxide-system-containing lozenge.BMC Microbiol. 2021 Nov 3;21(1):302. doi: 10.1186/s12866-021-02333-9. BMC Microbiol. 2021. PMID: 34732139 Free PMC article. Clinical Trial.
-
Antimicrobial photodynamic inactivation is potentiated by the addition of selenocyanate: Possible involvement of selenocyanogen?J Biophotonics. 2018 Aug;11(8):e201800029. doi: 10.1002/jbio.201800029. Epub 2018 May 6. J Biophotonics. 2018. PMID: 29488327 Free PMC article.
-
The Significance of Lactoperoxidase System in Oral Health: Application and Efficacy in Oral Hygiene Products.Int J Mol Sci. 2019 Mar 21;20(6):1443. doi: 10.3390/ijms20061443. Int J Mol Sci. 2019. PMID: 30901933 Free PMC article. Review.
-
Atmospheric Reactive Oxygen Species and Some Aspects of the Antiviral Protection at the Respiratory Epithelium.Biochem Mosc Suppl B Biomed Chem. 2022;16(2):79-90. doi: 10.1134/S1990750822020068. Epub 2022 May 17. Biochem Mosc Suppl B Biomed Chem. 2022. PMID: 35601461 Free PMC article.
References
-
- Zámocký M., Obinger C. Molecular phylogeny of heme peroxidases. In: Torres E., Ayala M., editors. Biocatalysis based on Heme Peroxidases. 1st ed. Springer; Berlin: 2010.
-
- Furtmuller P.G., Zederbauer M., Jantschko W. Active site structure and catalytic mechanisms of human peroxidases. Arch. Biochem. Biophys. 2006;445:199–213. - PubMed
-
- Arnhold J., Flemmig J. Human myeloperoxidase in innate and acquired immunity. Arch. Biochem. Biophys. 2010;500:92–106. - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
