

Phage mobility is a core determinant of phage-bacteria coexistence in biofilms
- PMID: 29125597
- PMCID: PMC5776469
- DOI: 10.1038/ismej.2017.190
Phage mobility is a core determinant of phage-bacteria coexistence in biofilms
Abstract
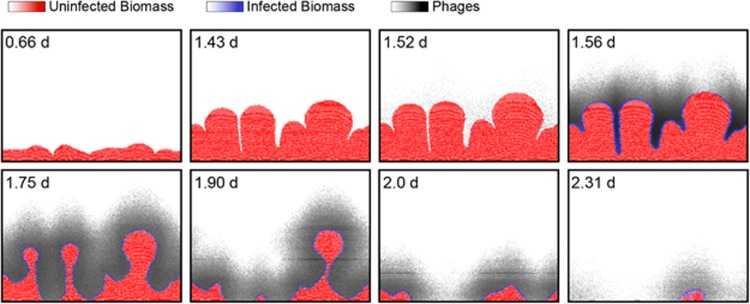
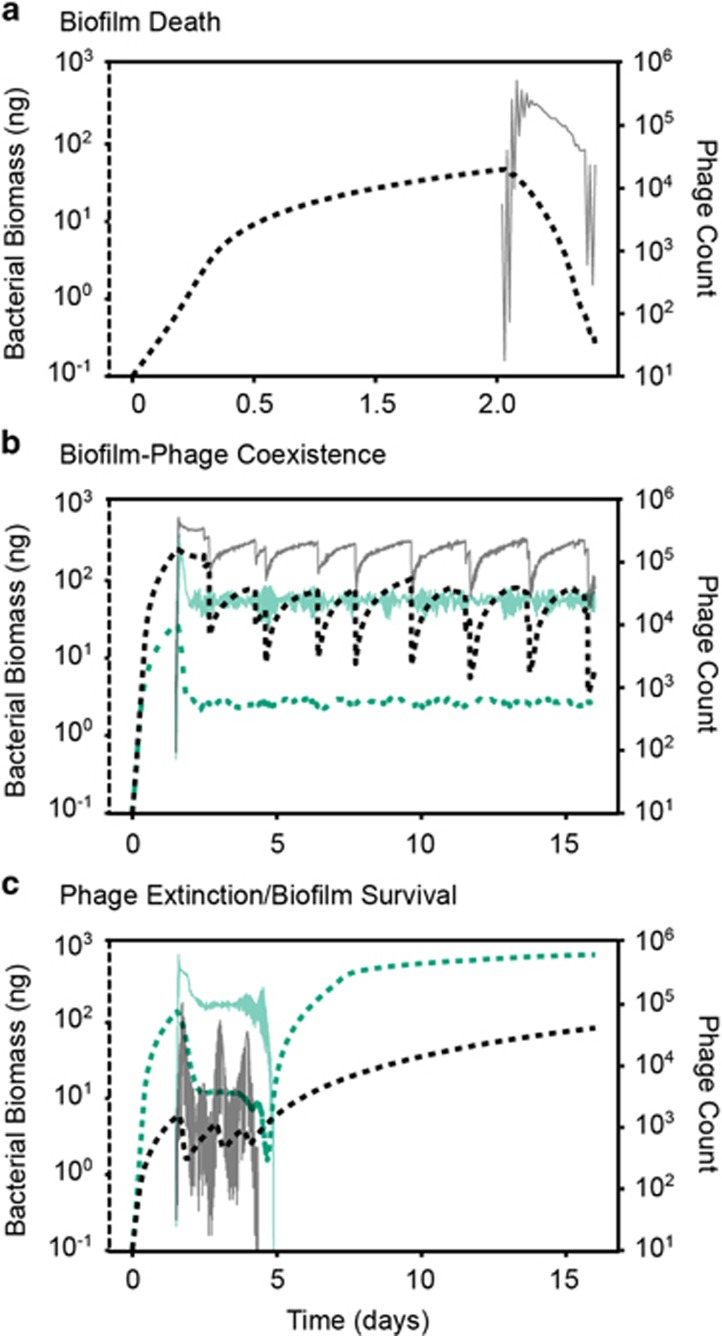
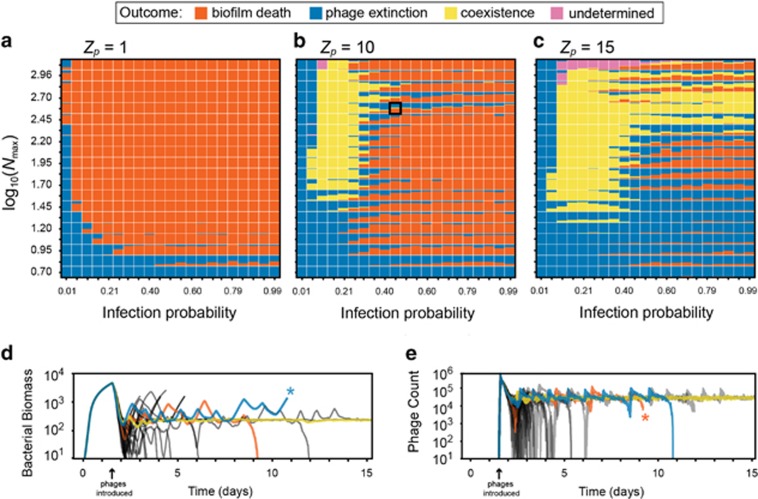
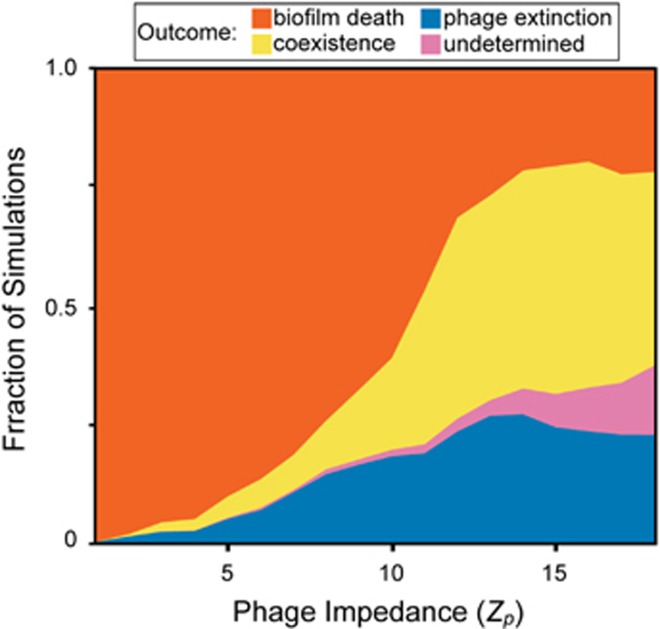
Many bacteria are adapted for attaching to surfaces and for building complex communities, termed biofilms. The biofilm mode of life is predominant in bacterial ecology. So too is the exposure of bacteria to ubiquitous viral pathogens, termed bacteriophages. Although biofilm-phage encounters are likely to be common in nature, little is known about how phages might interact with biofilm-dwelling bacteria. It is also unclear how the ecological dynamics of phages and their hosts depend on the biological and physical properties of the biofilm environment. To make headway in this area, we develop a biofilm simulation framework that captures key mechanistic features of biofilm growth and phage infection. Using these simulations, we find that the equilibrium state of interaction between biofilms and phages is governed largely by nutrient availability to biofilms, infection likelihood per host encounter and the ability of phages to diffuse through biofilm populations. Interactions between the biofilm matrix and phage particles are thus likely to be of fundamental importance, controlling the extent to which bacteria and phages can coexist in natural contexts. Our results open avenues to new questions of host-parasite coevolution and horizontal gene transfer in spatially structured biofilm contexts.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Abedon ST. (ed). (2008) Bacteriophage Ecology: Population Growth, Evolution, and Impact of Bacterial Viruses. Cambridge University Press: Cambridge, UK.
-
- Ackermann M. (2015). A functional perspective on phenotypic heterogeneity in microorganisms. Nat Rev Microbiol 13: 497–508. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
