

TM9SF4 is a novel factor promoting autophagic flux under amino acid starvation
- PMID: 29125601
- PMCID: PMC5762850
- DOI: 10.1038/cdd.2017.166
TM9SF4 is a novel factor promoting autophagic flux under amino acid starvation
Abstract
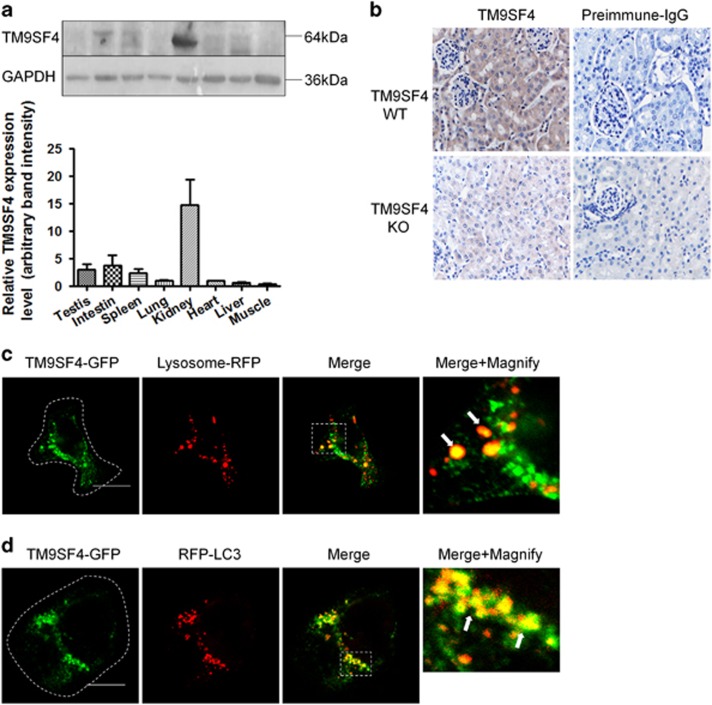
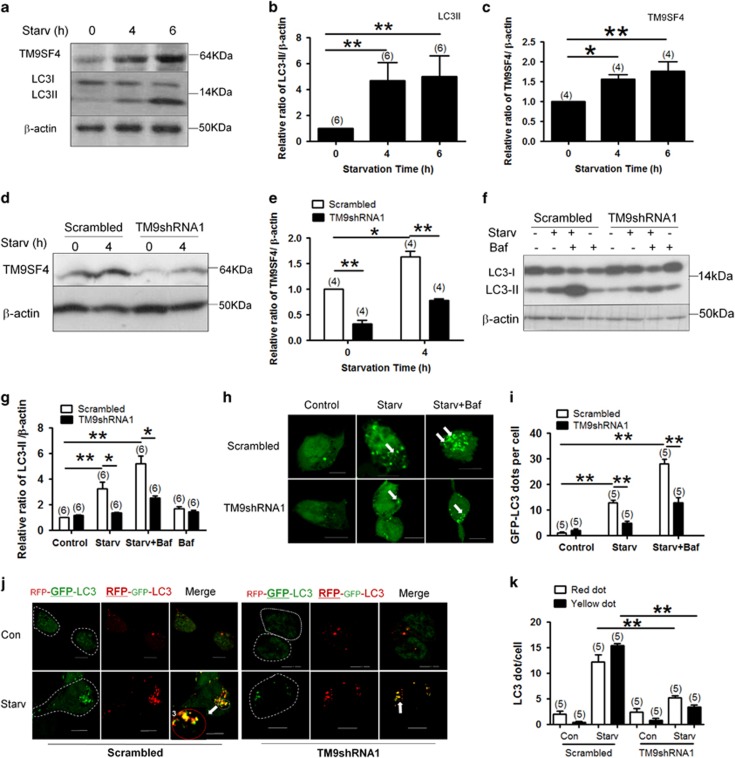
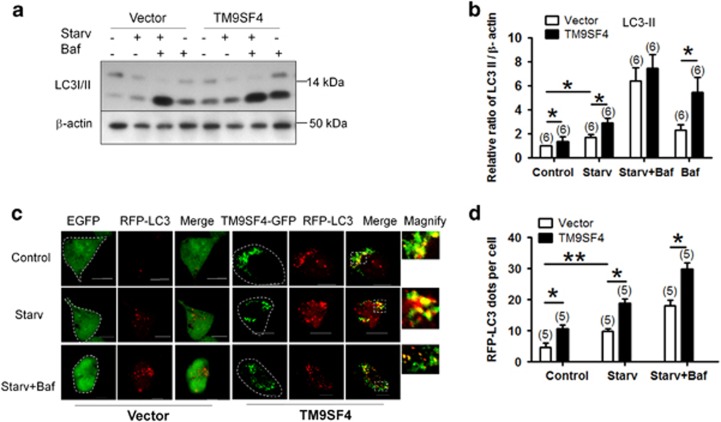
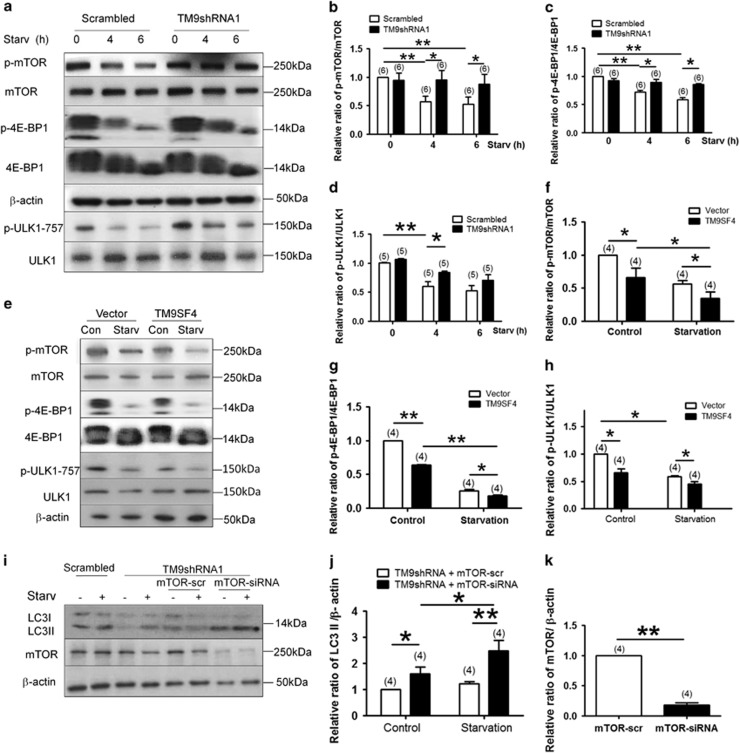
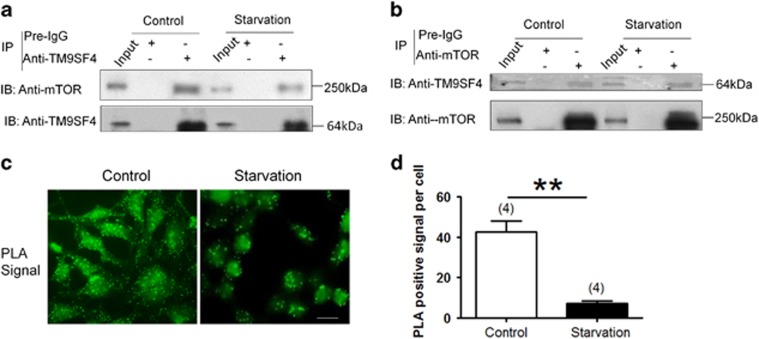
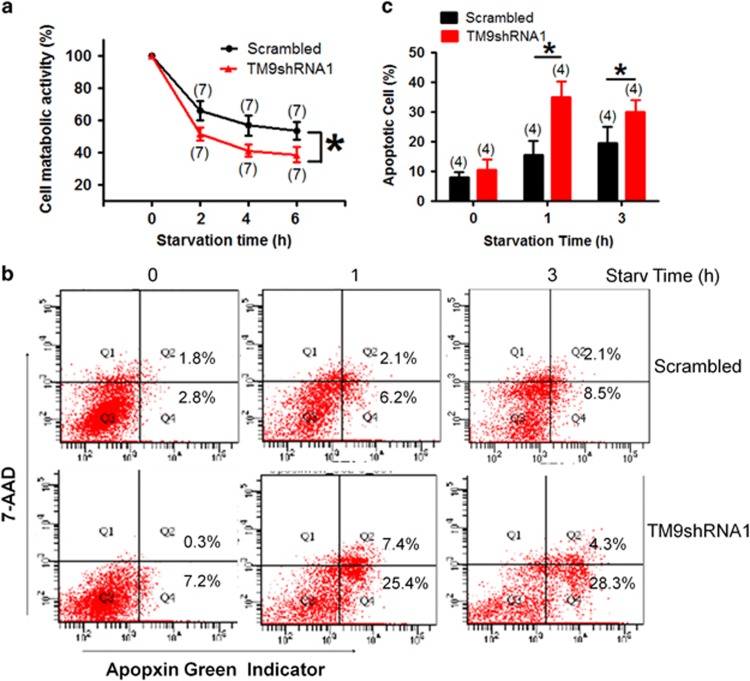
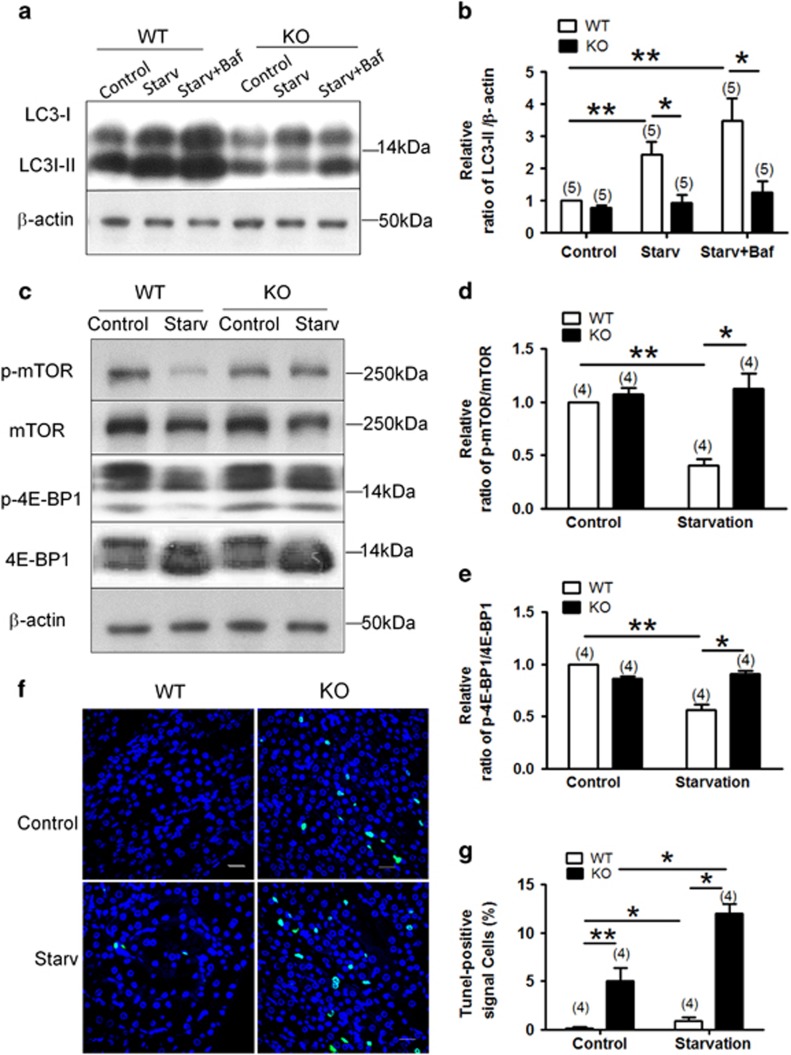
Autophagy is a highly complicated process with participation of large numbers of autophagy-related proteins. Under nutrient starvation, autophagy promotes cell survival by breaking down nonessential cellular components for recycling use. However, due to its high complexity, molecular mechanism of autophagy is still not fully understood. In the present study, we report a novel autophagy-related protein TM9SF4, which plays a functional role in the induction phase of autophagic process. TM9SF4 proteins were abundantly expressed in the kidney, especially in renal proximal tubular epithelial cells. At subcellular cells, TM9SF4 proteins were mostly localized in lysosome, Golgi, late endosome and autophagosome. Knockdown of TM9SF4 with TM9SF4-shRNAs markedly reduced the starvation-induced autophagy in HEK293 cells, the effect of which persisted in the presence of bafilomycin A1. TM9SF4-shRNAs also substantially attenuated the starvation-induced mTOR inactivation. In animal model, starvation was able to induce LC3-II accumulation and cause mTOR inactivation in renal cortical tissue in wild-type mice, the effect of which was minimal/absent in TM9SF4 knockout (TM9SF4-/-) mice. Co-immunoprecipitation and proximity ligation assay demonstrated physical interaction of TM9SF4 proteins with mTOR. In addition, knockdown or knockout of TM9SF4 reduced the starvation-induced cell death in HEK293 cells and animal model. Taken together, the present study identifies TM9SF4 as a novel autophagy-related protein. Under nutrient starvation, TM9SF4 functions to facilitate mTOR inactivation, resulting in an enhanced autophagic flux, which serves to protect cells from apoptotic cell death.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Polycystin-2 Plays an Essential Role in Glucose Starvation-Induced Autophagy in Human Embryonic Stem Cell-Derived Cardiomyocytes.Stem Cells. 2018 Apr;36(4):501-513. doi: 10.1002/stem.2764. Epub 2018 Jan 3. Stem Cells. 2018. PMID: 29271023
-
Endolysosomal two-pore channels regulate autophagy in cardiomyocytes.J Physiol. 2016 Jun 1;594(11):3061-77. doi: 10.1113/JP271332. Epub 2016 Feb 4. J Physiol. 2016. PMID: 26757341 Free PMC article.
-
TMEM74, a lysosome and autophagosome protein, regulates autophagy.Biochem Biophys Res Commun. 2008 May 2;369(2):622-9. doi: 10.1016/j.bbrc.2008.02.055. Epub 2008 Feb 21. Biochem Biophys Res Commun. 2008. PMID: 18294959
-
AMPK Inhibits ULK1-Dependent Autophagosome Formation and Lysosomal Acidification via Distinct Mechanisms.Mol Cell Biol. 2018 Apr 30;38(10):e00023-18. doi: 10.1128/MCB.00023-18. Print 2018 May 15. Mol Cell Biol. 2018. PMID: 29507183 Free PMC article.
-
Glucose induces autophagy under starvation conditions by a p38 MAPK-dependent pathway.Biochem J. 2013 Jan 15;449(2):497-506. doi: 10.1042/BJ20121122. Biochem J. 2013. PMID: 23116132
Cited by
-
A Golgi-Located Transmembrane Nine Protein Gene TMN11 Functions in Manganese/Cadmium Homeostasis and Regulates Growth and Seed Development in Rice.Int J Mol Sci. 2022 Dec 14;23(24):15883. doi: 10.3390/ijms232415883. Int J Mol Sci. 2022. PMID: 36555524 Free PMC article.
-
Classification of Cell-in-Cell Structures: Different Phenomena with Similar Appearance.Cells. 2021 Sep 28;10(10):2569. doi: 10.3390/cells10102569. Cells. 2021. PMID: 34685548 Free PMC article. Review.
-
Mechanisms and significance of entosis for tumour growth and progression.Cell Death Discov. 2024 Mar 1;10(1):109. doi: 10.1038/s41420-024-01877-9. Cell Death Discov. 2024. PMID: 38429285 Free PMC article. Review.
-
Life, death and autophagy.Nat Cell Biol. 2018 Oct;20(10):1110-1117. doi: 10.1038/s41556-018-0201-5. Epub 2018 Sep 17. Nat Cell Biol. 2018. PMID: 30224761 Free PMC article. Review.
-
TRPM2 promotes autophagic degradation in vascular smooth muscle cells.Sci Rep. 2020 Nov 26;10(1):20719. doi: 10.1038/s41598-020-77620-y. Sci Rep. 2020. PMID: 33244095 Free PMC article.
References
-
- Chluba-de Tapia J, de Tapia M, Jaggin V, Eberle AN. Cloning of a human multispanning membrane protein cDNA: evidence for a new protein family. Gene 1997; 197: 195–204. - PubMed
-
- Fais S, Fauvarque MO. TM9 and cannibalism: how to learn more about cancer by studying amoebae and invertebrates. Trends Mol Med 2012; 18: 4–5. - PubMed
-
- Froquet R, Cherix N, Birke R, Benghezal M, Cameroni E, Letourneur F et al. Control of cellular physiology by TM9 proteins in yeast and Dictyostelium. J Biol Chem 2008; 283: 6764–6772. - PubMed
-
- Gao C, Cai Y, Wang Y, Kang BH, Aniento F, Robinson DG et al. Retention mechanisms for ER and Golgi membrane proteins. Trends Plant Sci 2014; 19: 508–515. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
