

A Chemoproteomic Approach to Query the Degradable Kinome Using a Multi-kinase Degrader
- PMID: 29129717
- PMCID: PMC6427047
- DOI: 10.1016/j.chembiol.2017.10.005
A Chemoproteomic Approach to Query the Degradable Kinome Using a Multi-kinase Degrader
Abstract
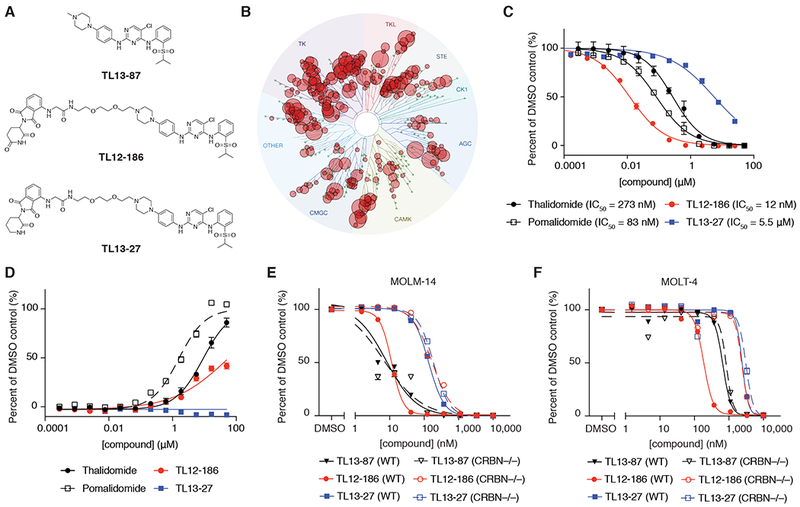
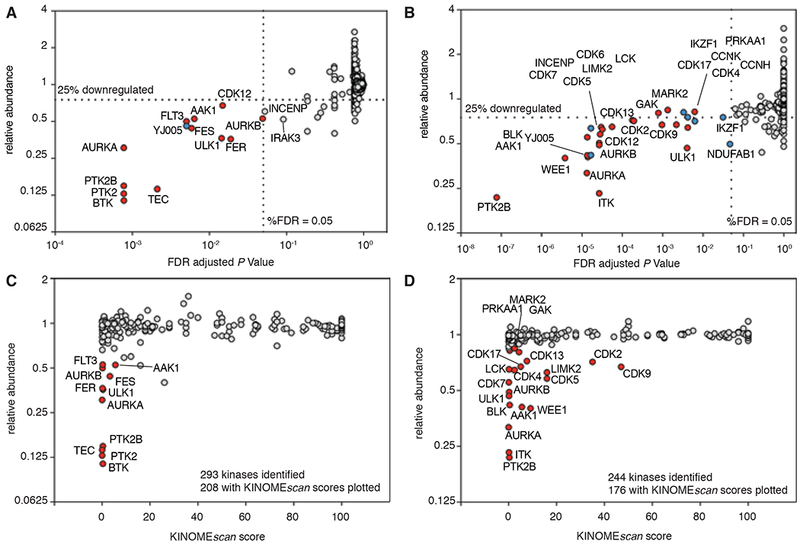
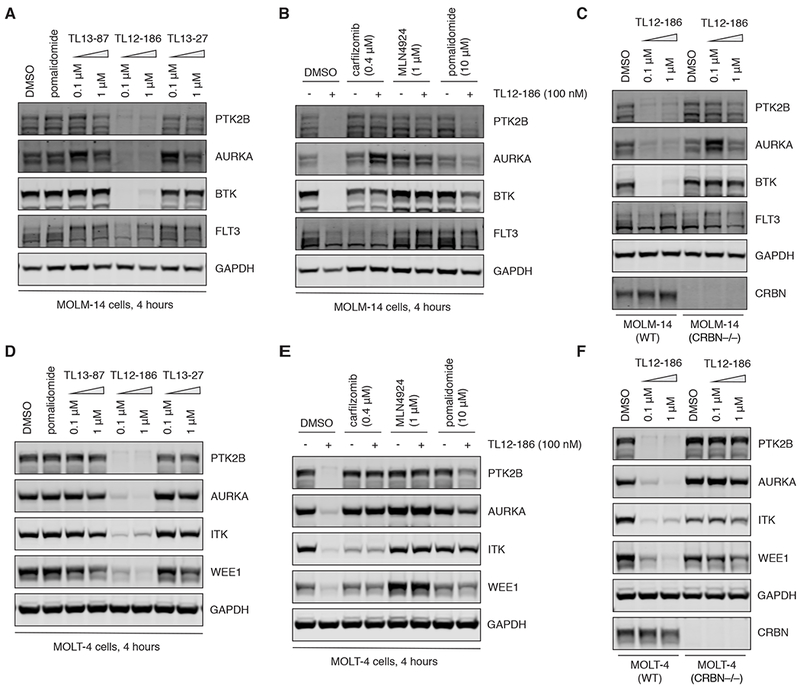
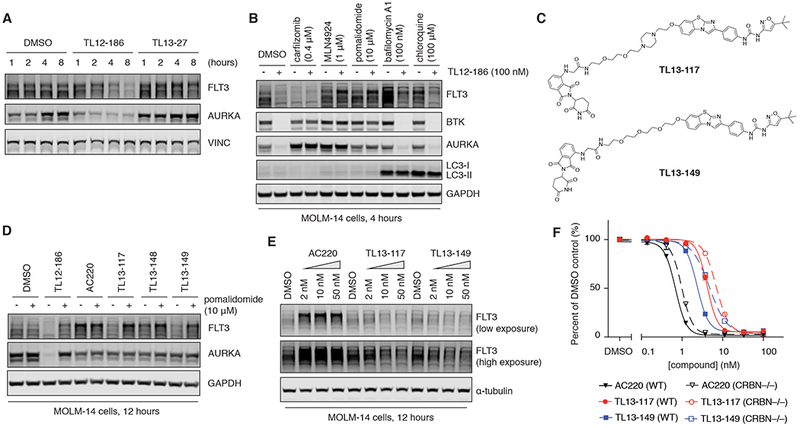
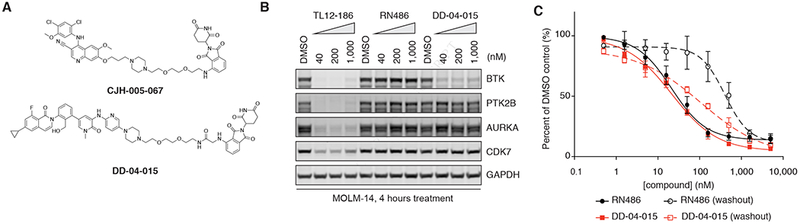
Heterobifunctional molecules that recruit E3 ubiquitin ligases, such as cereblon, for targeted protein degradation represent an emerging pharmacological strategy. A major unanswered question is how generally applicable this strategy is to all protein targets. In this study, we designed a multi-kinase degrader by conjugating a highly promiscuous kinase inhibitor with a cereblon-binding ligand, and used quantitative proteomics to discover 28 kinases, including BTK, PTK2, PTK2B, FLT3, AURKA, AURKB, TEC, ULK1, ITK, and nine members of the CDK family, as degradable. This set of kinases is only a fraction of the intracellular targets bound by the degrader, demonstrating that successful degradation requires more than target engagement. The results guided us to develop selective degraders for FLT3 and BTK, with potentials to improve disease treatment. Together, this study demonstrates an efficient approach to triage a gene family of interest to identify readily degradable targets for further studies and pre-clinical developments.
Keywords: BTK; FLT3; chemoproteomics; drug design; kinase; protein degradation.
Copyright © 2017 Elsevier Ltd. All rights reserved.
Figures
Comment in
-
Try Me: Promiscuous Inhibitors Still Allow for Selective Targeted Protein Degradation.Cell Chem Biol. 2018 Jan 18;25(1):4-6. doi: 10.1016/j.chembiol.2018.01.004. Cell Chem Biol. 2018. PMID: 29351837
References
-
- Bacher U, Haferlach C, Kern W, Haferlach T, and Schnittger S (2008). Prognostic relevance of FLT3-TKD mutations in AML: the combination matters--an analysis of 3082 patients. Blood 111, 2527–2537. - PubMed
-
- Boschelli DH, Ye F, Wang YD, Dutia M, Johnson SL, Wu B, Miller K, Powell DW, Yaczko D, Young M, et al. (2001). Optimization of 4-phenylamino-3-quinolinecarbonitriles as potent inhibitors of Src kinase activity. J Med Chem 44, 3965–3977. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
