

Amphiphysin (BIN1) negatively regulates dynamin 2 for normal muscle maturation
- PMID: 29130937
- PMCID: PMC5707152
- DOI: 10.1172/JCI90542
Amphiphysin (BIN1) negatively regulates dynamin 2 for normal muscle maturation
Abstract
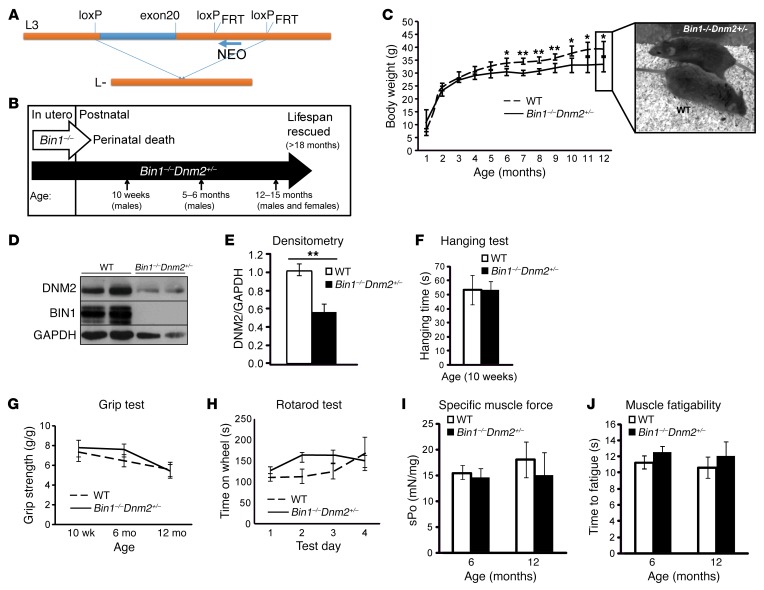
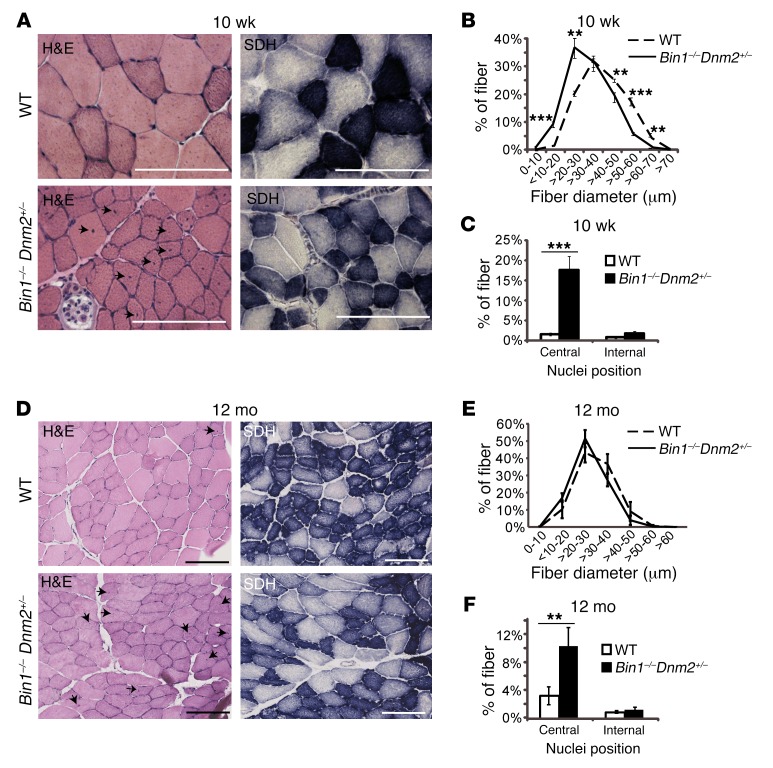
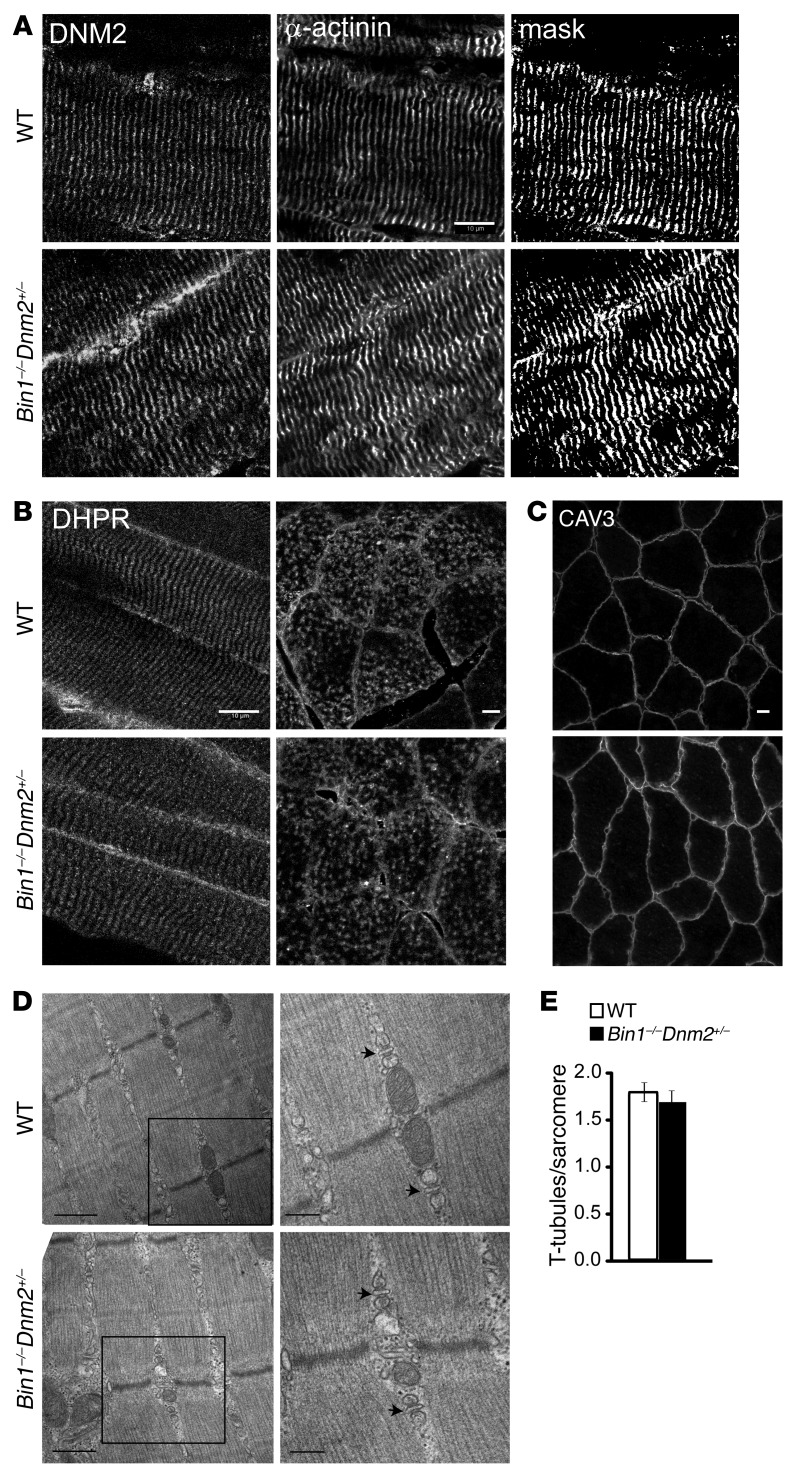
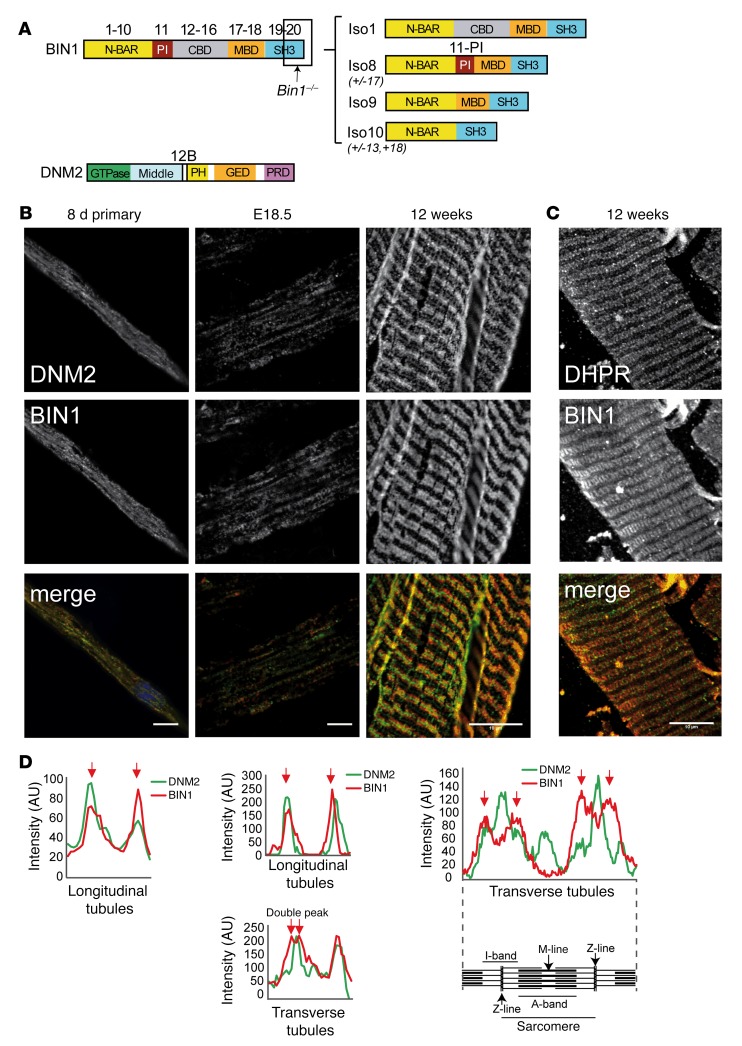
Regulation of skeletal muscle development and organization is a complex process that is not fully understood. Here, we focused on amphiphysin 2 (BIN1, also known as bridging integrator-1) and dynamin 2 (DNM2), two ubiquitous proteins implicated in membrane remodeling and mutated in centronuclear myopathies (CNMs). We generated Bin1-/- Dnm2+/- mice to decipher the physiological interplay between BIN1 and DNM2. While Bin1-/- mice die perinatally from a skeletal muscle defect, Bin1-/- Dnm2+/- mice survived at least 18 months, and had normal muscle force and intracellular organization of muscle fibers, supporting BIN1 as a negative regulator of DNM2. We next characterized muscle-specific isoforms of BIN1 and DNM2. While BIN1 colocalized with and partially inhibited DNM2 activity during muscle maturation, BIN1 had no effect on the isoform of DNM2 found in adult muscle. Together, these results indicate that BIN1 and DNM2 regulate muscle development and organization, function through a common pathway, and define BIN1 as a negative regulator of DNM2 in vitro and in vivo during muscle maturation. Our data suggest that DNM2 modulation has potential as a therapeutic approach for patients with CNM and BIN1 defects. As BIN1 is implicated in cancers, arrhythmia, and late-onset Alzheimer disease, these findings may trigger research directions and therapeutic development for these common diseases.
Keywords: Muscle Biology; Neuromuscular disease; Skeletal muscle.
Conflict of interest statement
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
