

Small RNA teg49 Is Derived from a sarA Transcript and Regulates Virulence Genes Independent of SarA in Staphylococcus aureus
- PMID: 29133345
- PMCID: PMC5778362
- DOI: 10.1128/IAI.00635-17
Small RNA teg49 Is Derived from a sarA Transcript and Regulates Virulence Genes Independent of SarA in Staphylococcus aureus
Abstract
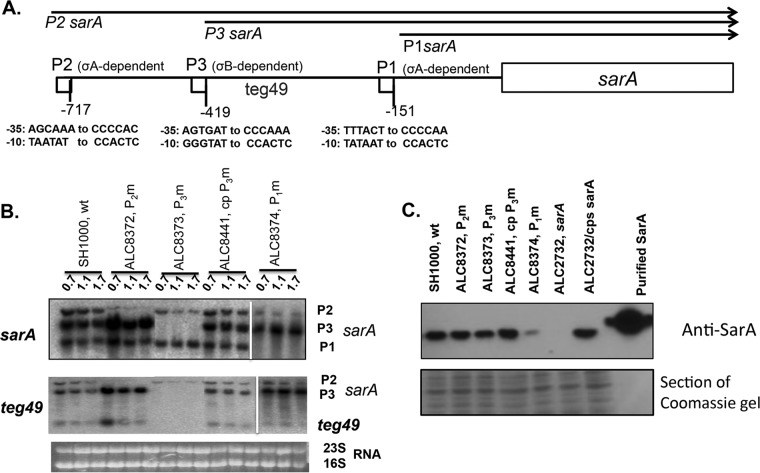
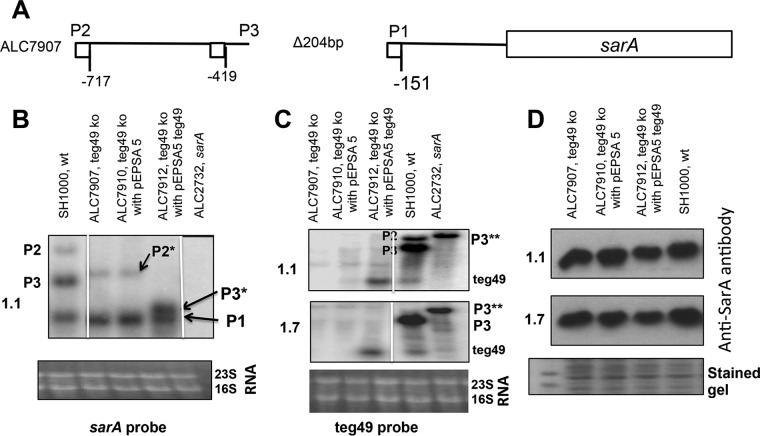
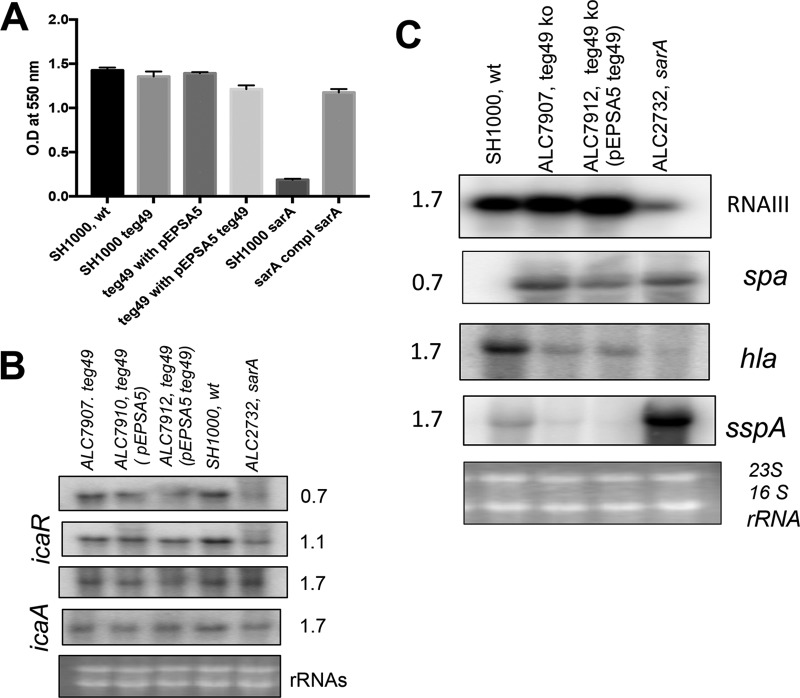
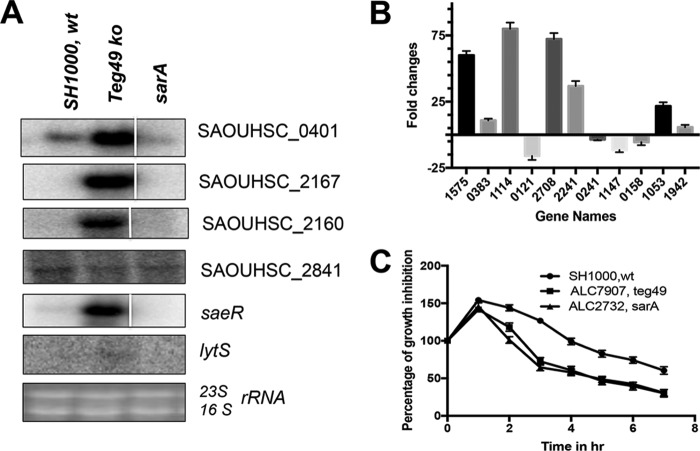
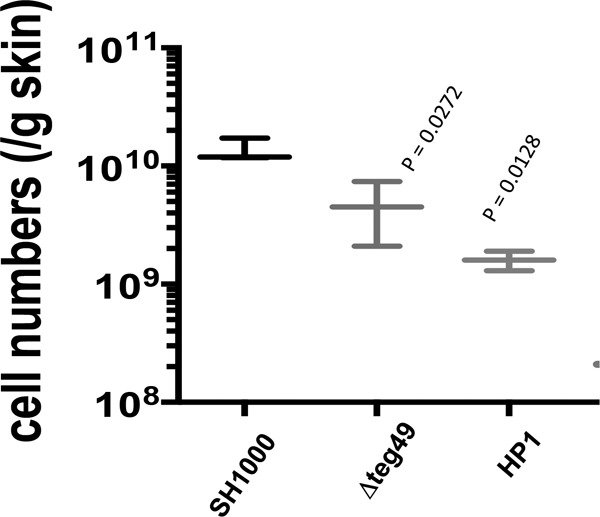
Expression of virulence factors in Staphylococcus aureus is regulated by a wide range of transcriptional regulators, including proteins and small RNAs (sRNAs), at the level of transcription and/or translation. The sarA locus consists of three overlapping transcripts generated from three distinct promoters, all containing the sarA open reading frame (ORF). The 5' untranslated regions (UTRs) of these transcripts contain three separate regions ∼711, 409, and 146 nucleotides (nt) upstream of the sarA translation start, the functions of which remain unknown. Recent transcriptome-sequencing (RNA-Seq) analysis and subsequent characterization indicated that two sRNAs, teg49 and teg48, are processed and likely produced from the sarA P3 and sarA P1 transcripts of the sarA locus, respectively. In this report, we utilized a variety of sarA promoter mutants and cshA and rnc mutants to ascertain the contributions of these factors to the generation of teg49. We also defined the transcriptional regulon of teg49, including virulence genes not regulated by SarA. Phenotypically, teg49 did not impact biofilm formation or affect overall SarA expression significantly. Comparative analyses of RNA-Seq data between the wild-type, teg49 mutant, and sarA mutant strains indicated that ∼133 genes are significantly upregulated while 97 are downregulated in a teg49 deletion mutant in a sarA-independent manner. An abscess model of skin infection indicated that the teg49 mutant exhibited a reduced bacterial load compared to the wild-type S. aureus Overall, these results suggest that teg49 sRNA has a regulatory role in target gene regulation independent of SarA. The exact mechanism of this regulation is yet to be dissected.
Keywords: RNA-Seq; S. aureus; SarA; biofilms; gene expression; gene regulation; mouse model; small RNAs.
Copyright © 2018 American Society for Microbiology.
Figures

References
-
- Crossley KB, Archer GL. 1997. The staphylococci in human disease. Churchill Livingston, New York, NY.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
