

Role of Calprotectin in Withholding Zinc and Copper from Candida albicans
- PMID: 29133349
- PMCID: PMC5778358
- DOI: 10.1128/IAI.00779-17
Role of Calprotectin in Withholding Zinc and Copper from Candida albicans
Abstract
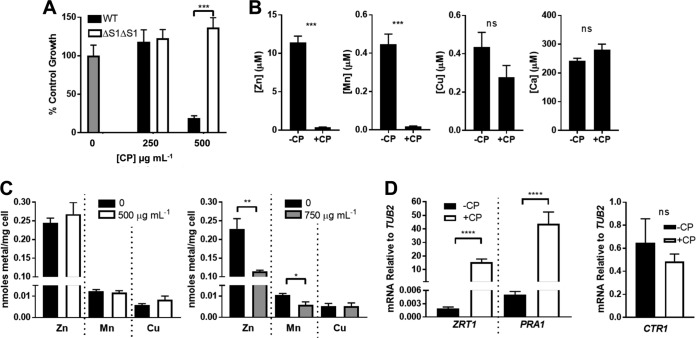
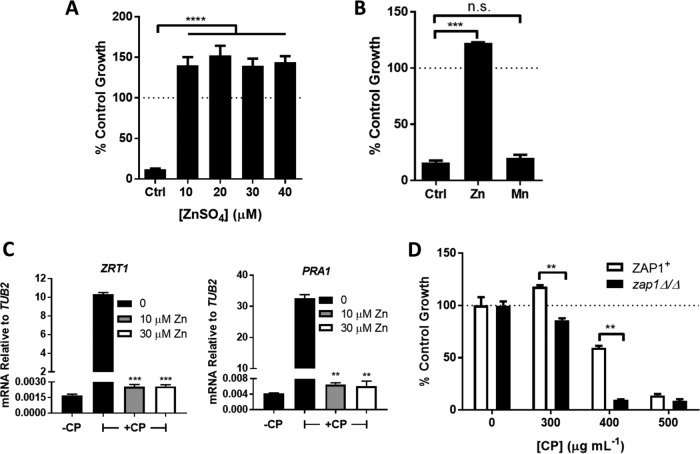
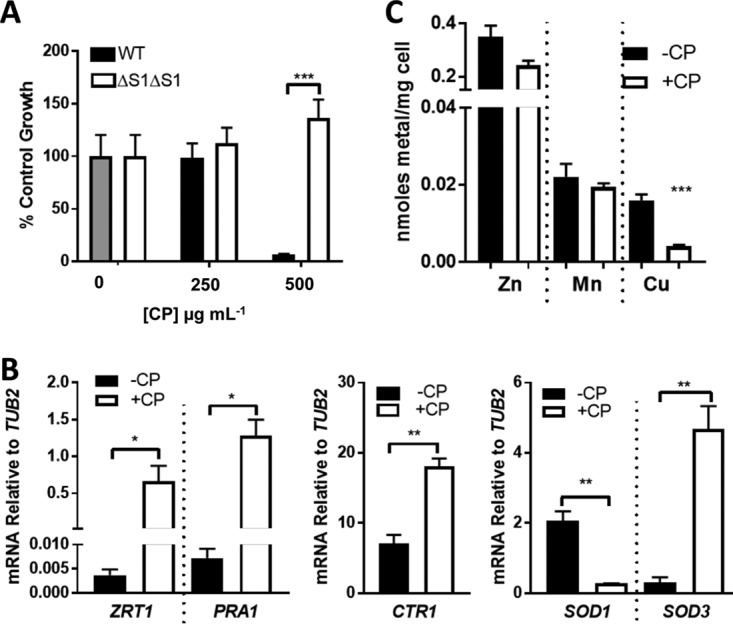
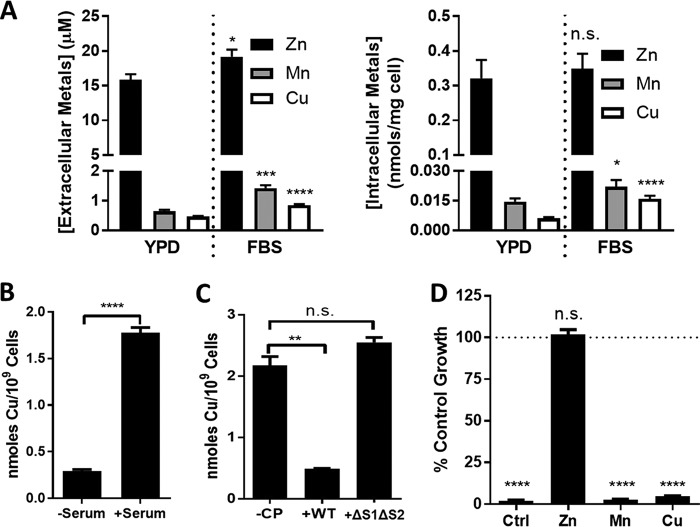
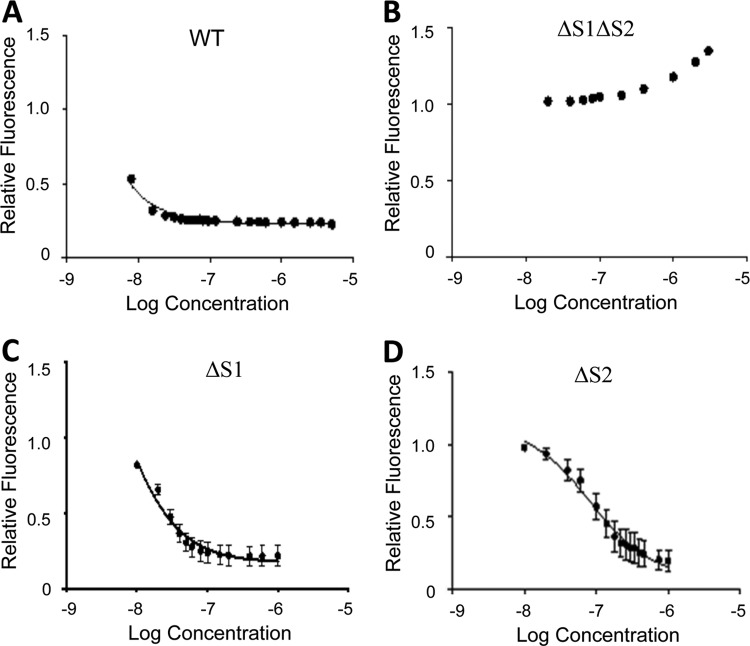
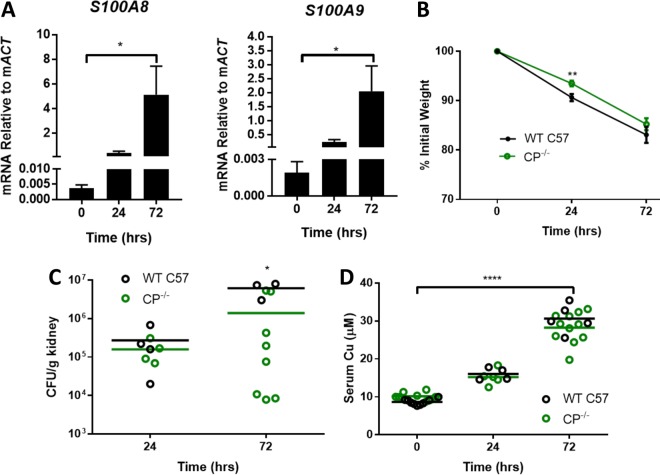
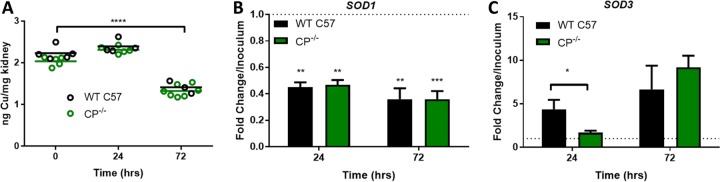
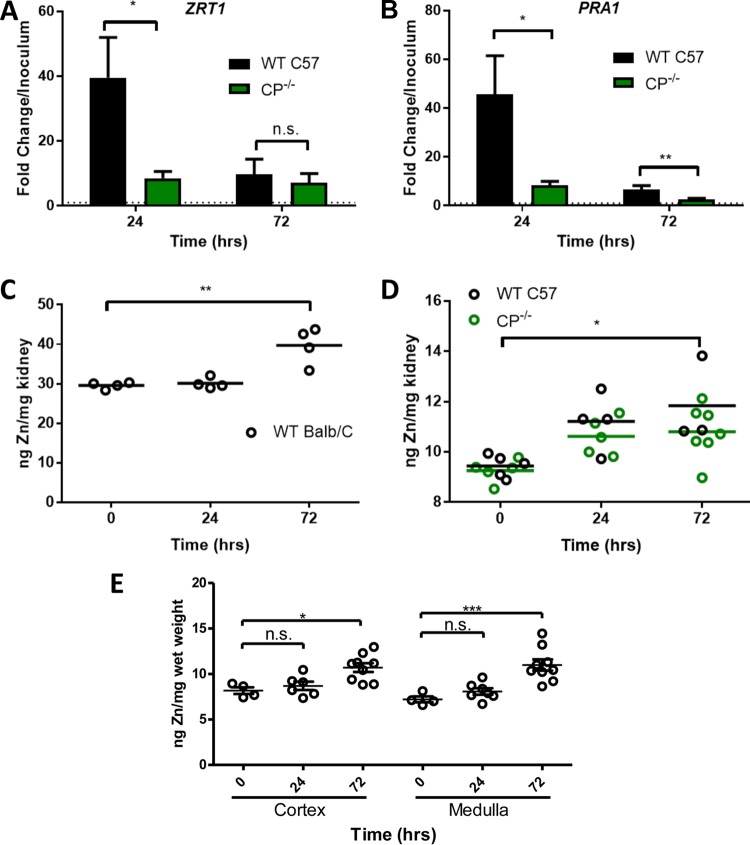
The opportunistic fungal pathogen Candida albicans acquires essential metals from the host, yet the host can sequester these micronutrients through a process known as nutritional immunity. How the host withholds metals from C. albicans has been poorly understood; here we examine the role of calprotectin (CP), a transition metal binding protein. When CP depletes bioavailable Zn from the extracellular environment, C. albicans strongly upregulates ZRT1 and PRA1 for Zn import and maintains constant intracellular Zn through numerous cell divisions. We show for the first time that CP can also sequester Cu by binding Cu(II) with subpicomolar affinity. CP blocks fungal acquisition of Cu from serum and induces a Cu starvation stress response involving SOD1 and SOD3 superoxide dismutases. These transcriptional changes are mirrored when C. albicans invades kidneys in a mouse model of disseminated candidiasis, although the responses to Cu and Zn limitations are temporally distinct. The Cu response progresses throughout 72 h, while the Zn response is short-lived. Notably, these stress responses were attenuated in CP null mice, but only at initial stages of infection. Thus, Zn and Cu pools are dynamic at the host-pathogen interface and CP acts early in infection to restrict metal nutrients from C. albicans.
Keywords: Candida albicans; calprotectin; copper; nutritional immunity; zinc.
Copyright © 2018 American Society for Microbiology.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
