

Correlating kinetic and structural data on ubiquinone binding and reduction by respiratory complex I
- PMID: 29133414
- PMCID: PMC5715780
- DOI: 10.1073/pnas.1714074114
Correlating kinetic and structural data on ubiquinone binding and reduction by respiratory complex I
Abstract
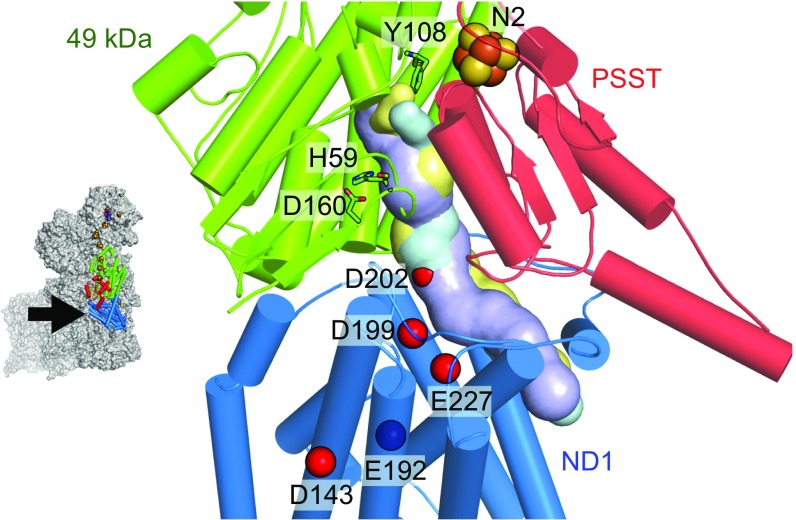
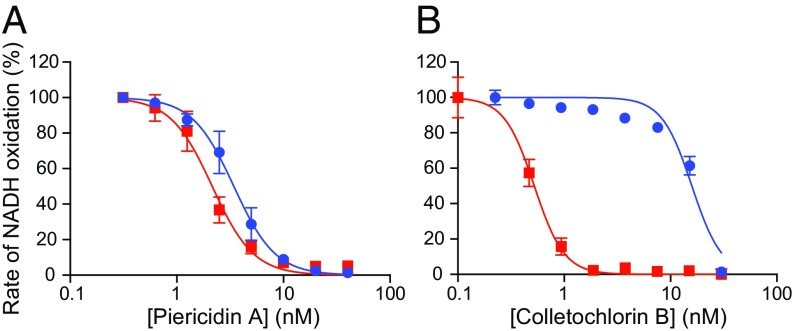
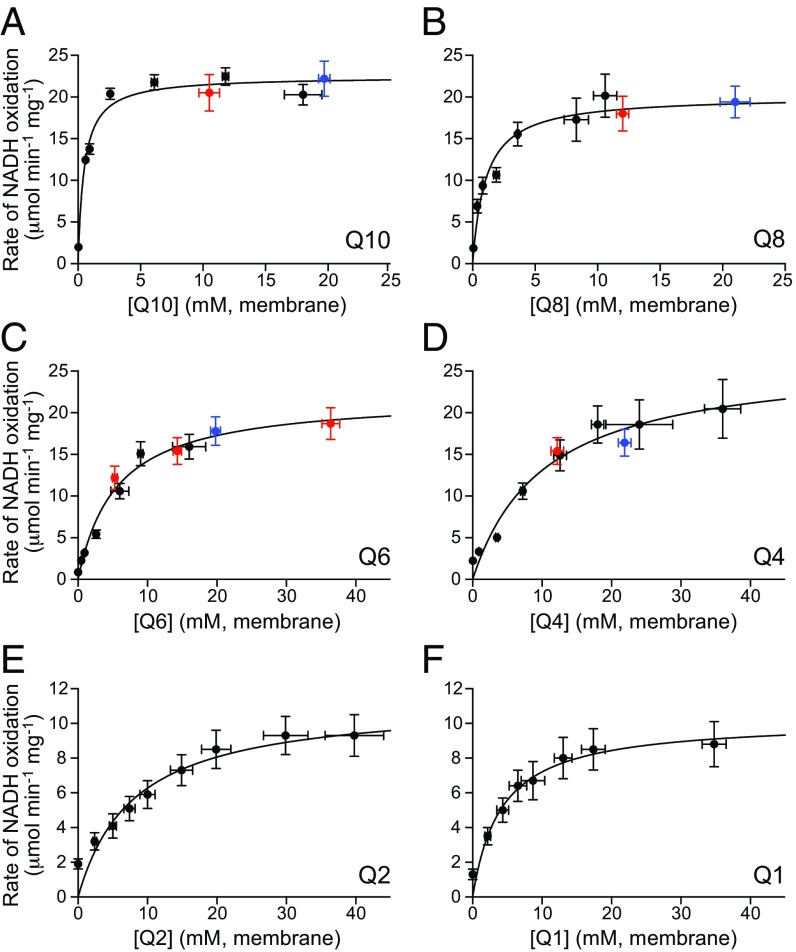
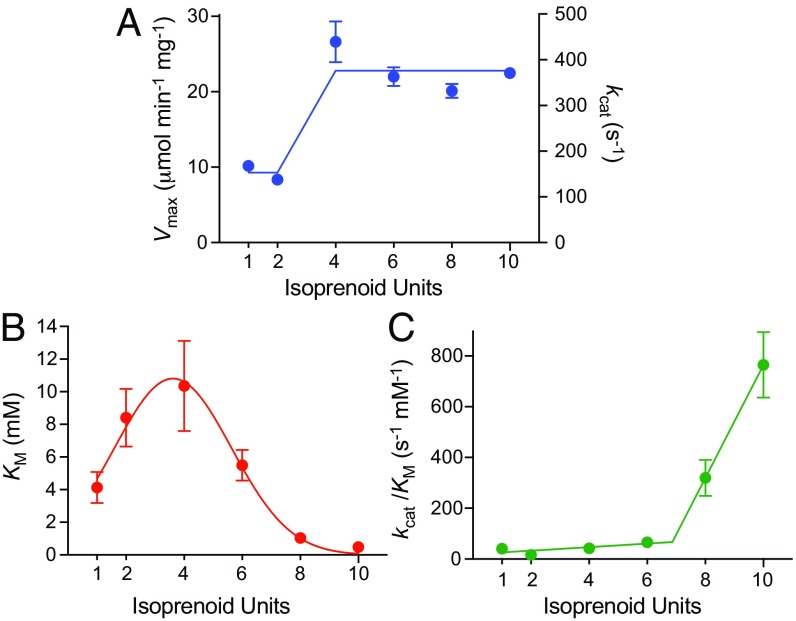
Respiratory complex I (NADH:ubiquinone oxidoreductase), one of the largest membrane-bound enzymes in mammalian cells, powers ATP synthesis by using the energy from electron transfer from NADH to ubiquinone-10 to drive protons across the energy-transducing mitochondrial inner membrane. Ubiquinone-10 is extremely hydrophobic, but in complex I the binding site for its redox-active quinone headgroup is ∼20 Å above the membrane surface. Structural data suggest it accesses the site by a narrow channel, long enough to accommodate almost all of its ∼50-Å isoprenoid chain. However, how ubiquinone/ubiquinol exchange occurs on catalytically relevant timescales, and whether binding/dissociation events are involved in coupling electron transfer to proton translocation, are unknown. Here, we use proteoliposomes containing complex I, together with a quinol oxidase, to determine the kinetics of complex I catalysis with ubiquinones of varying isoprenoid chain length, from 1 to 10 units. We interpret our results using structural data, which show the hydrophobic channel is interrupted by a highly charged region at isoprenoids 4-7. We demonstrate that ubiquinol-10 dissociation is not rate determining and deduce that ubiquinone-10 has both the highest binding affinity and the fastest binding rate. We propose that the charged region and chain directionality assist product dissociation, and that isoprenoid stepping ensures short transit times. These properties of the channel do not benefit the exhange of short-chain quinones, for which product dissociation may become rate limiting. Thus, we discuss how the long channel does not hinder catalysis under physiological conditions and the possible roles of ubiquinone/ubiquinol binding/dissociation in energy conversion.
Keywords: NADH:ubiquinone oxidoreductase; bioenergetics; coenzyme Q10; electron transport chain; mitochondria.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
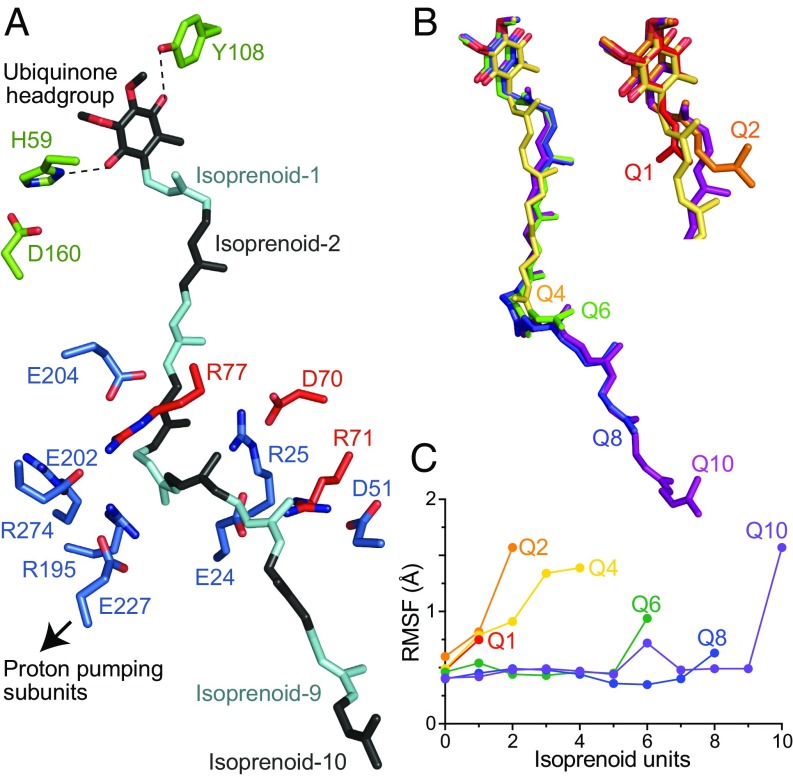
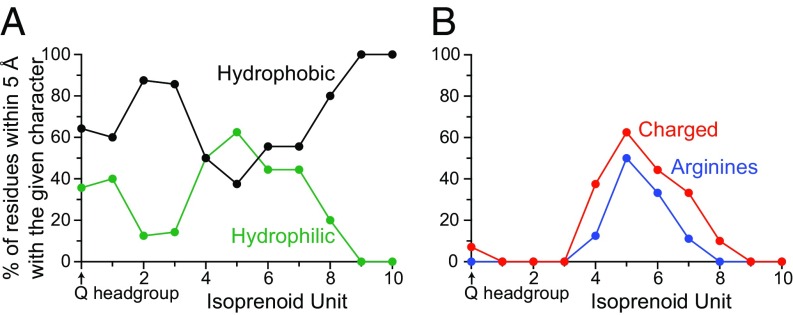
References
-
- Hirst J. Mitochondrial complex I. Annu Rev Biochem. 2013;82:551–575. - PubMed
-
- Galkin AS, Grivennikova VG, Vinogradov AD. →H+/2e- stoichiometry in NADH-quinone reductase reactions catalyzed by bovine heart submitochondrial particles. FEBS Lett. 1999;451:157–161. - PubMed
-
- Fassone E, Rahman S. Complex I deficiency: Clinical features, biochemistry and molecular genetics. J Med Genet. 2012;49:578–590. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
