

Early Postnatal Cardiomyocyte Proliferation Requires High Oxidative Energy Metabolism
- PMID: 29133820
- PMCID: PMC5684334
- DOI: 10.1038/s41598-017-15656-3
Early Postnatal Cardiomyocyte Proliferation Requires High Oxidative Energy Metabolism
Abstract
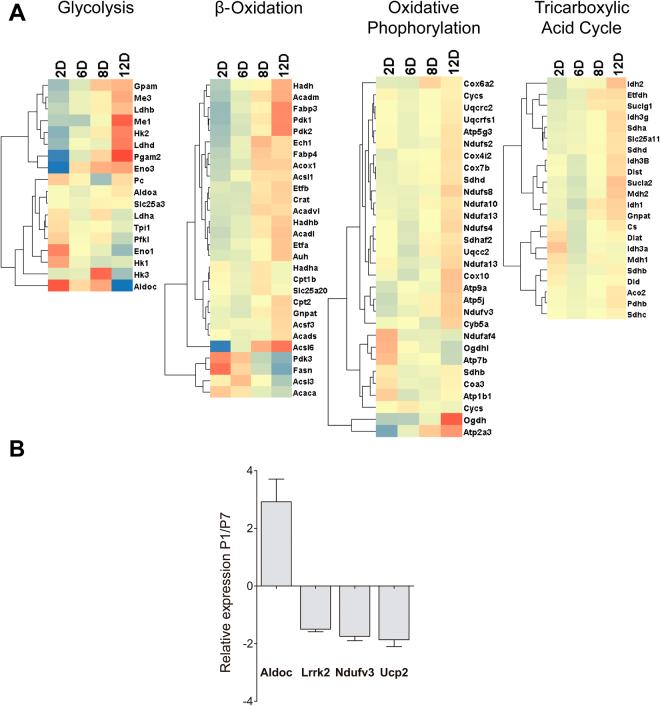
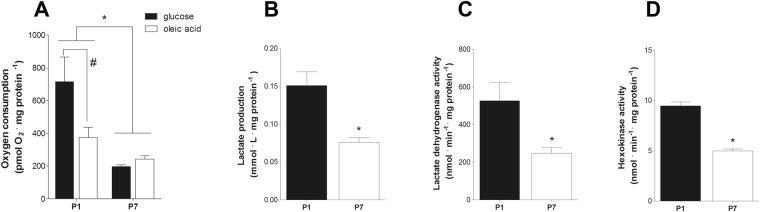
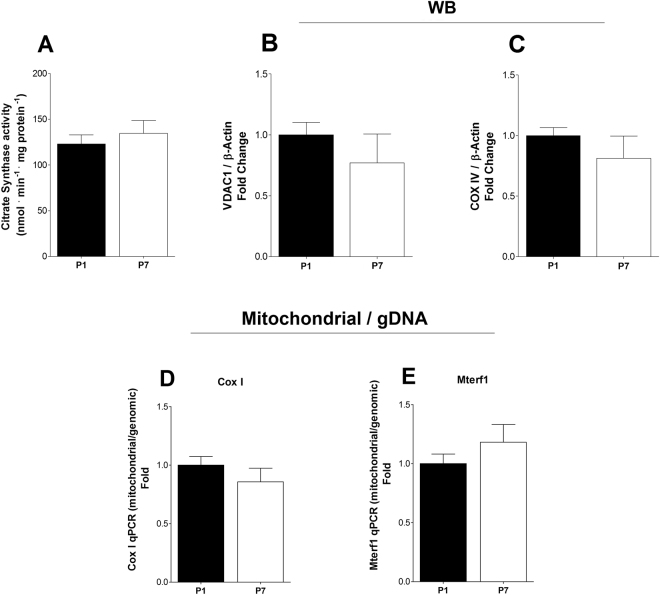
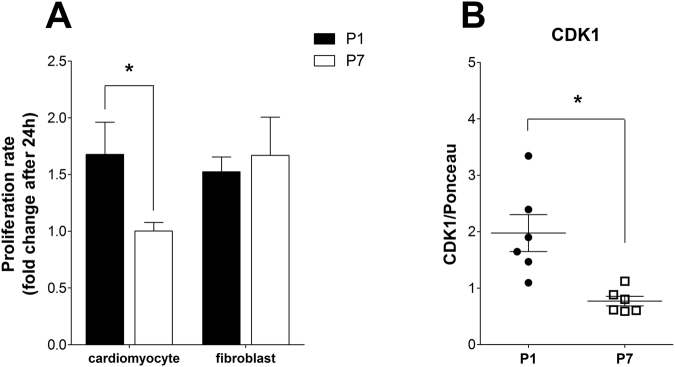
Cardiac energy metabolism must cope with early postnatal changes in tissue oxygen tensions, hemodynamics, and cell proliferation to sustain development. Here, we tested the hypothesis that proliferating neonatal cardiomyocytes are dependent on high oxidative energy metabolism. We show that energy-related gene expression does not correlate with functional oxidative measurements in the developing heart. Gene expression analysis suggests a gradual overall upregulation of oxidative-related genes and pathways, whereas functional assessment in both cardiac tissue and cultured cardiomyocytes indicated that oxidative metabolism decreases between the first and seventh days after birth. Cardiomyocyte extracellular flux analysis indicated that the decrease in oxidative metabolism between the first and seventh days after birth was mostly related to lower rates of ATP-linked mitochondrial respiration, suggesting that overall energetic demands decrease during this period. In parallel, the proliferation rate was higher for early cardiomyocytes. Furthermore, in vitro nonlethal chemical inhibition of mitochondrial respiration reduced the proliferative capacity of early cardiomyocytes, indicating a high energy demand to sustain cardiomyocyte proliferation. Altogether, we provide evidence that early postnatal cardiomyocyte proliferative capacity correlates with high oxidative energy metabolism. The energy requirement decreases as the proliferation ceases in the following days, and both oxidative-dependent metabolism and anaerobic glycolysis subside.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures



References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
