

A free-boundary model of a motile cell explains turning behavior
- PMID: 29136638
- PMCID: PMC5705165
- DOI: 10.1371/journal.pcbi.1005862
A free-boundary model of a motile cell explains turning behavior
Abstract
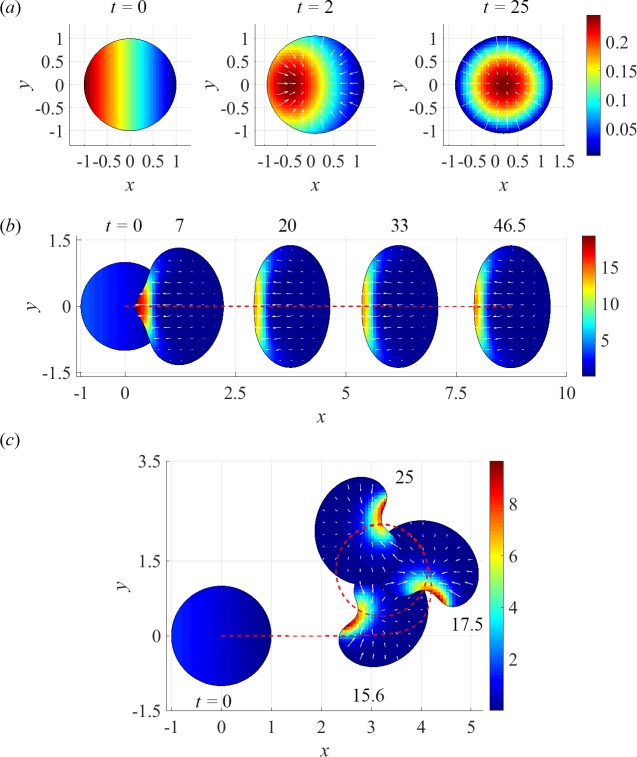
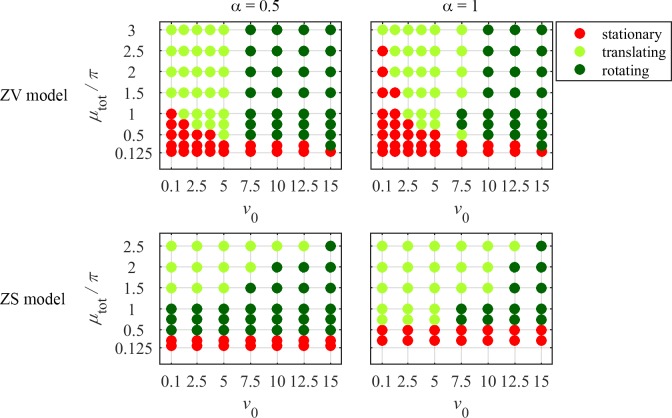
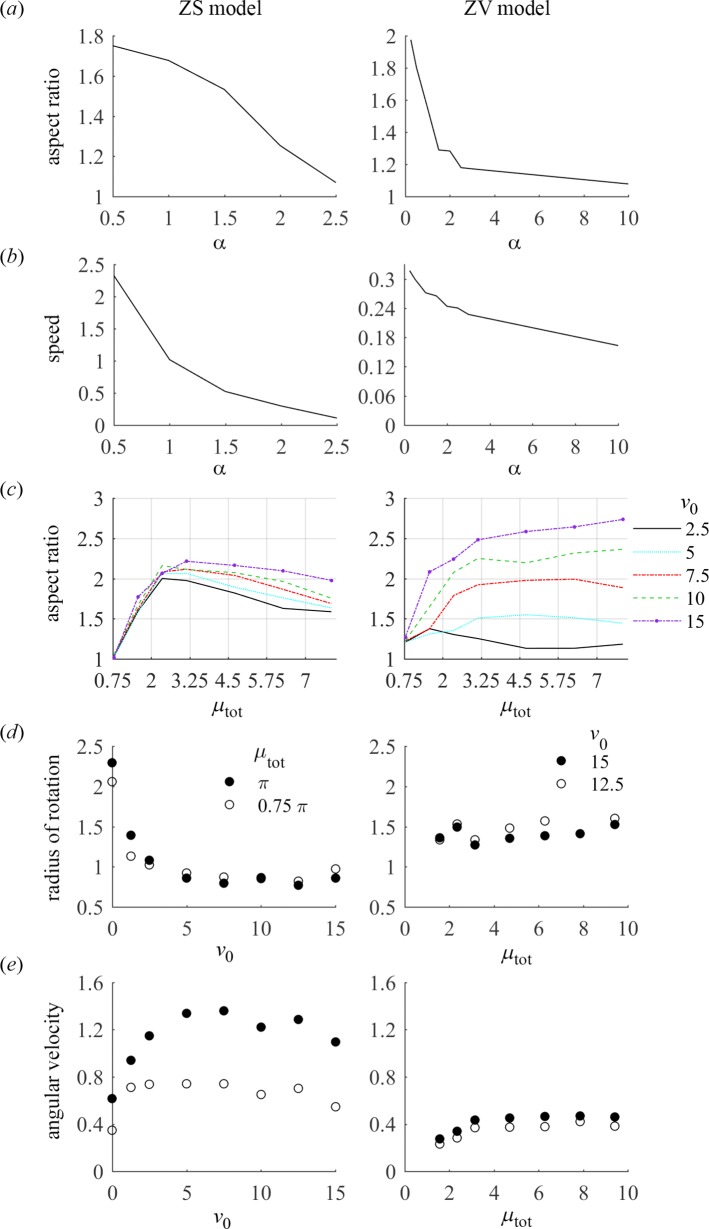
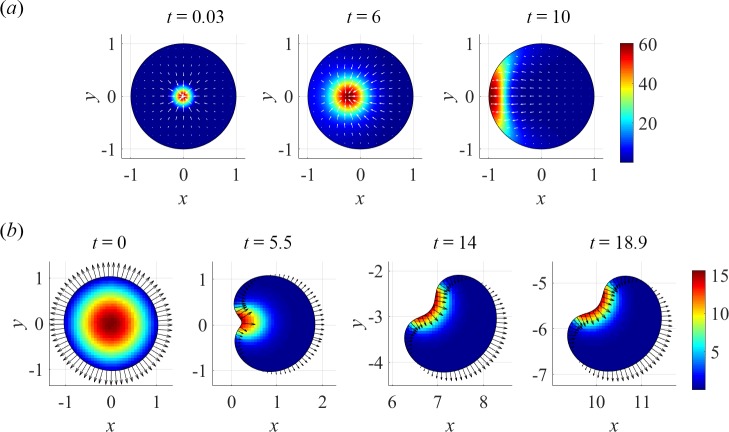
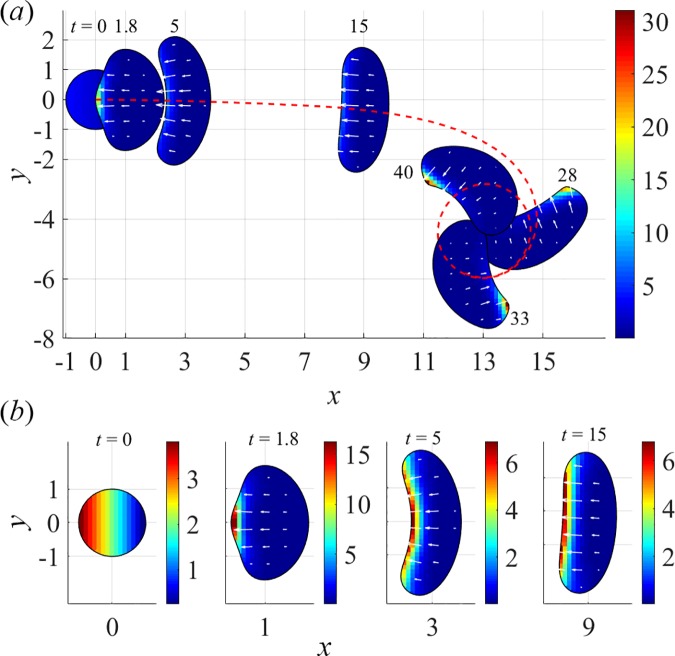
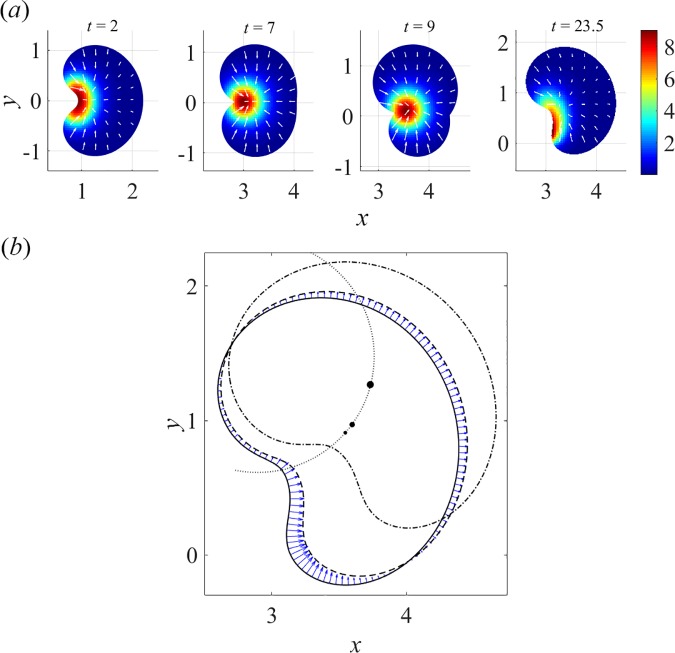
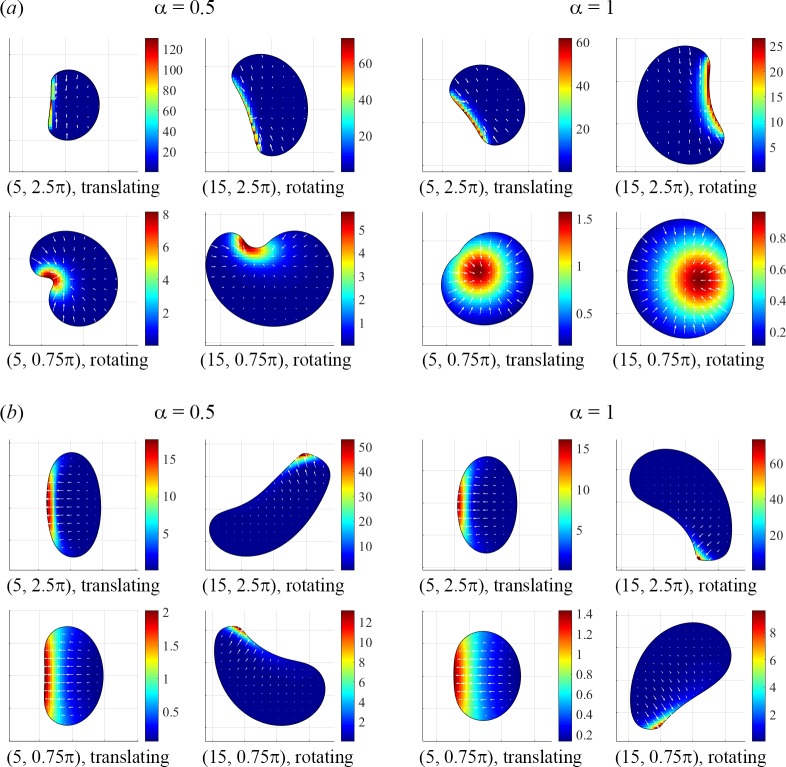
To understand shapes and movements of cells undergoing lamellipodial motility, we systematically explore minimal free-boundary models of actin-myosin contractility consisting of the force-balance and myosin transport equations. The models account for isotropic contraction proportional to myosin density, viscous stresses in the actin network, and constant-strength viscous-like adhesion. The contraction generates a spatially graded centripetal actin flow, which in turn reinforces the contraction via myosin redistribution and causes retraction of the lamellipodial boundary. Actin protrusion at the boundary counters the retraction, and the balance of the protrusion and retraction shapes the lamellipodium. The model analysis shows that initiation of motility critically depends on three dimensionless parameter combinations, which represent myosin-dependent contractility, a characteristic viscosity-adhesion length, and a rate of actin protrusion. When the contractility is sufficiently strong, cells break symmetry and move steadily along either straight or circular trajectories, and the motile behavior is sensitive to conditions at the cell boundary. Scanning of a model parameter space shows that the contractile mechanism of motility supports robust cell turning in conditions where short viscosity-adhesion lengths and fast protrusion cause an accumulation of myosin in a small region at the cell rear, destabilizing the axial symmetry of a moving cell.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures

References
-
- Ridley AJ, Schwartz MA, Burridge K, Firtel RA, Ginsberg MH, Borisy G, et al. Cell migration: integrating signals from front to back. Science 2003. December 5;302(5651):1704–9. doi: 10.1126/science.1092053 - DOI - PubMed
-
- Lämmermann T, Sixt M. Mechanical modes of 'amoeboid' cell migration. Curr Opin Cell Biol. 2009. October;21(5):636–44. doi: 10.1016/j.ceb.2009.05.003 - DOI - PubMed
-
- Verkhovsky AB. The mechanisms of spatial and temporal patterning of cell-edge dynamics. Curr Opin Cell Biol. 2015. October;36: 113–21. doi: 10.1016/j.ceb.2015.09.001 - DOI - PubMed
-
- Keren K, Pincus Z, Allen GM, Barnhart EL, Marriott G, Mogilner A, et al. Mechanism of shape determination in motile cells. Nature 2008. May 22;453(7194):475–80. doi: 10.1038/nature06952 - DOI - PMC - PubMed
-
- Barnhart EL, Lee KC, Keren K, Mogilner A, Theriot JA. An adhesion-dependent switch between mechanisms that determine motile cell shape. PLoS Biol. 2011. May;9(5): e1001059 doi: 10.1371/journal.pbio.1001059 - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
