

Regulation of FBXO4-mediated ICAM-1 protein stability in metastatic breast cancer
- PMID: 29137327
- PMCID: PMC5669953
- DOI: 10.18632/oncotarget.20912
Regulation of FBXO4-mediated ICAM-1 protein stability in metastatic breast cancer
Abstract
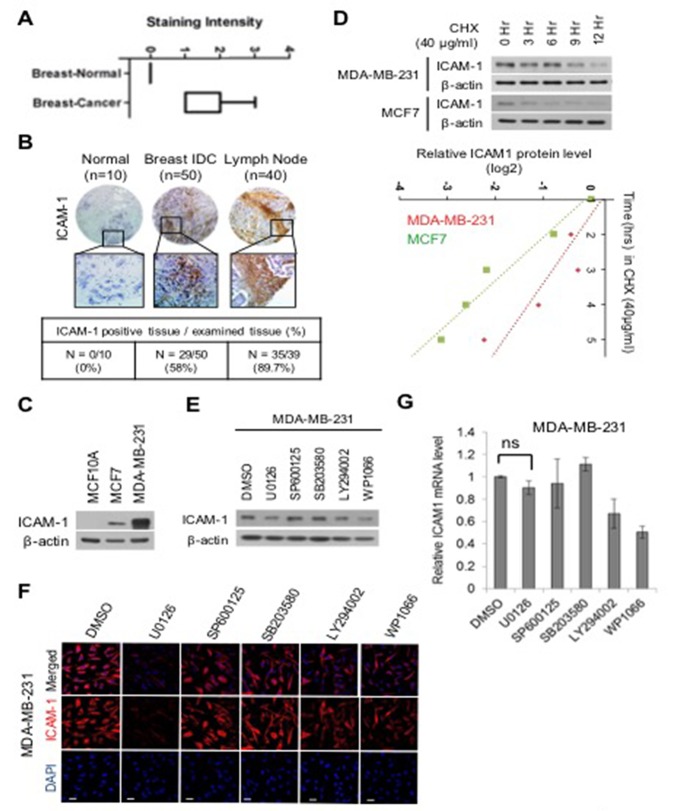
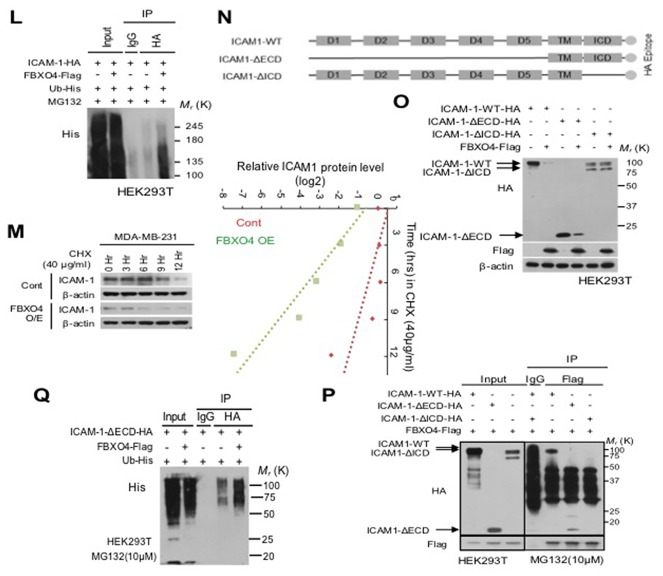
Advanced or progressive cancers share common traits such as altered transcriptional modulation, genetic modification, and abnormal post-translational regulation. These processes influence protein stability and cellular activity. Intercellular adhesion molecule-1 (ICAM-1) is involved in the malignant progression of various human cancers, including breast, liver, renal, and pancreatic cancers, but protein stability has not been deal with in metastatic breast cancer. Additionally, the relevance of the stability maintenance of ICAM-1 protein remains obscure. Here, we identified a novel interaction of E3 ligase FBXO4 that is specifically presented to ICAM-1. To understand how FBXO4 modulates ICAM-1 stability, we investigated ICAM-1-overexpressing or knockdown metastatic/non-metastatic breast cancers. ICAM-1 was found to influence tumor progression and metastasis, whereas FBXO4 regulated aggressive tumorigenic conditions. These results demonstrate that FBXO4 is a major regulator of ICAM-1 stability and that alterations in the stability of ICAM-1 can influence therapeutic outcome in metastatic cancer.
Keywords: E3 ligase; FBXO4; intercellular adhesion molecule-1; metastatic breast cancer; stability.
Conflict of interest statement
CONFLICTS OF INTEREST The authors declare that they have no conflicts of interest.
Figures




References
-
- Hanahan D, Weinberg RA. Hallmarks of cancer: the next generation. Cell. 2011;144:646–74. - PubMed
-
- Badve S, Dabbs DJ, Schnitt SJ, Baehner FL, Decker T, Eusebi V, Fox SB, Ichihara S, Jacquemier J, Lakhani SR, Palacios J, Rakha EA, Richardson AL, et al. Basal-like and triple-negative breast cancers: a critical review with an emphasis on the implications for pathologists and oncologists. Mod Pathol. 2011;24:157–67. - PubMed
-
- Duperray A, Languino LR, Plescia J, McDowall A, Hogg N, Craig AG, Berendt AR, Altieri DC. Molecular identification of a novel fibrinogen binding site on the first domain of ICAM-1 regulating leukocyte-endothelium bridging. J Biol Chem. 1997;272:435–41. - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
