

Downregulation of the Sonic Hedgehog/Gli pathway transcriptional target Neogenin-1 is associated with basal cell carcinoma aggressiveness
- PMID: 29137400
- PMCID: PMC5663572
- DOI: 10.18632/oncotarget.21061
Downregulation of the Sonic Hedgehog/Gli pathway transcriptional target Neogenin-1 is associated with basal cell carcinoma aggressiveness
Abstract
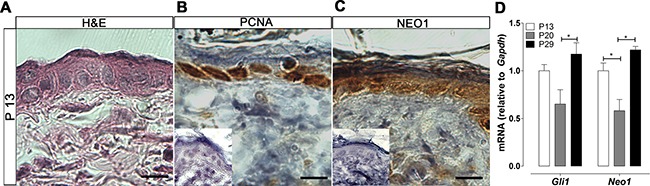
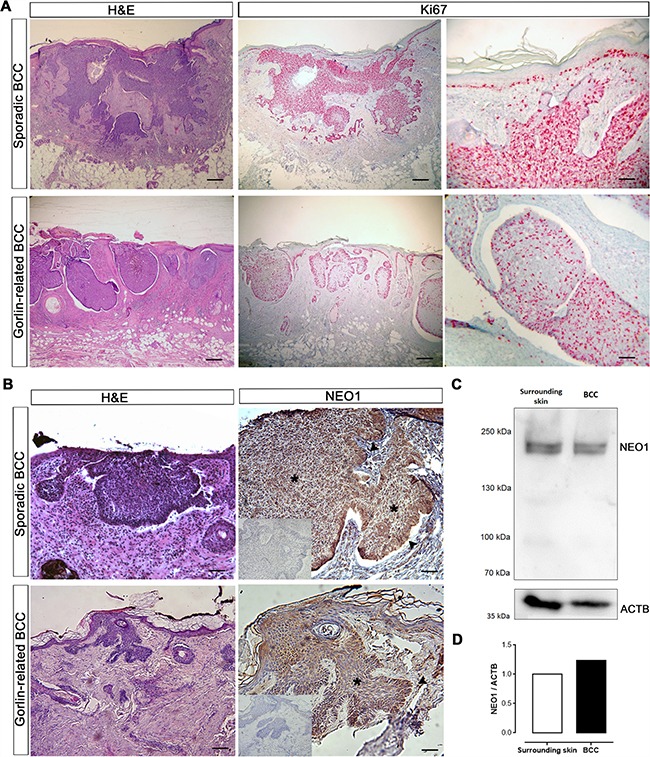
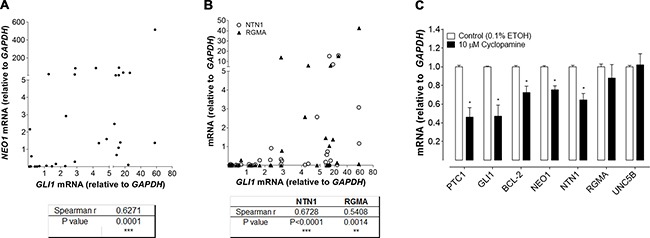
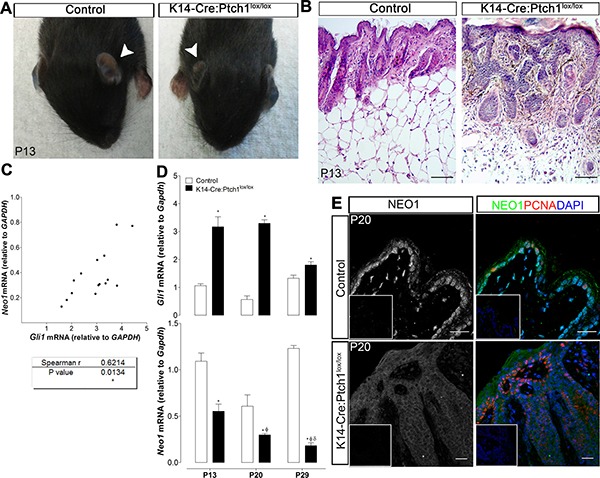
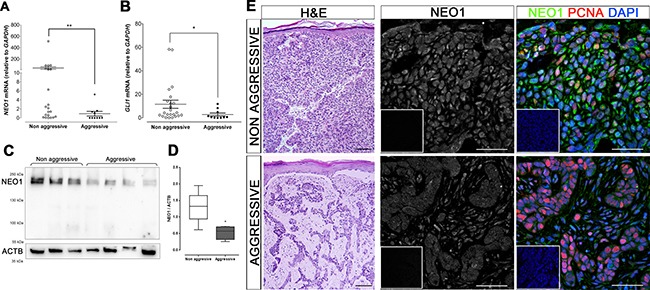
Basal Cell Carcinoma (BCC) is one of the most diagnosed cancers worldwide. It develops due to an unrestrained Sonic Hedgehog (SHH) signaling activity in basal cells of the skin. Certain subtypes of BCC are more aggressive than others, although the molecular basis of this phenomenon remains unknown. We have previously reported that Neogenin-1 (NEO1) is a downstream target gene of the SHH/GLI pathway in neural tissue. Given that SHH participates in epidermal homeostasis, here we analyzed the epidermal expression of NEO1 in order to identify whether it plays a role in adult epidermis or BCC. We describe the mRNA and protein expression profile of NEO1 and its ligands (Netrin-1 and RGMA) in human and mouse control epidermis and in a broad range of human BCCs. We identify in human BCC a significant positive correlation in the levels of NEO1 receptor, NTN-1 and RGMA ligands with respect to GLI1, the main target gene of the canonical SHH pathway. Moreover, we show via cyclopamine inhibition of the SHH/GLI pathway of ex vivo cultures that NEO1 likely functions as a downstream target of SHH/GLI signaling in the skin. We also show how Neo1 expression decreases throughout BCC progression in the K14-Cre:Ptch1lox/lox mouse model and that aggressive subtypes of human BCC exhibit lower levels of NEO1 than non-aggressive BCC samples. Taken together, these data suggest that NEO1 is a SHH/GLI target in epidermis. We propose that NEO1 may be important in tumor onset and is then down-regulated in advanced BCC or aggressive subtypes.
Keywords: BCC; Neogenin-1; Netrin-1; SHH/GLI pathway; tumor aggressiveness.
Conflict of interest statement
CONFLICTS OF INTEREST The authors of this paper declare no conflicts of interest.
Figures

References
-
- Madan V, Lear JT, Szeimies R. Non-melanoma skin cancer. Lancet. 2010;375:673–85. https://doi.org/10.1016/S0140-6736(09)61196-X. - DOI - PubMed
-
- Aszterbaum M, Epstein J, Oro A, Douglas V, LeBoit PE, Scott MP, Epstein EH. Ultraviolet and ionizing radiation enhance the growth of BCCs and trichoblastomas in patched heterozygous knockout mice. Nat Med. 1999;5:1285–91. https://doi.org/10.1038/15242. - DOI - PubMed
-
- Yu M, Zloty D, Cowan B, Shapiro J, Haegert A, Bell RH, Warshawski L, Carr N, McElwee KJ. Superficial, nodular, and morpheiform basal-cell carcinomas exhibit distinct gene expression profiles. J Invest Dermatol. 2008;128:1797–805. https://doi.org/10.1038/sj.jid.5701243. - DOI - PubMed
-
- Kasper M, Jaks V, Hohl D, Toftgård R. Review series basal cell carcinoma: molecular biology and potential new therapies. J Clin Invest. 2012;122:455–63. https://doi.org/10.1172/JCI58779. - DOI - PMC - PubMed
-
- Crowson AN. Basal cell carcinoma: biology, morphology, and clinical implications. Mod Pathol. 2006;19:S127–47. https://doi.org/10.1038/modpathol.3800512. - DOI - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
