

The photocycle of orange carotenoid protein conceals distinct intermediates and asynchronous changes in the carotenoid and protein components
- PMID: 29138423
- PMCID: PMC5686206
- DOI: 10.1038/s41598-017-15520-4
The photocycle of orange carotenoid protein conceals distinct intermediates and asynchronous changes in the carotenoid and protein components
Abstract
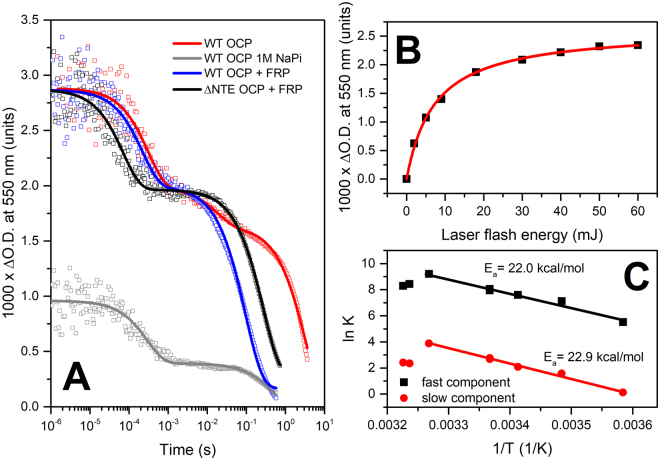
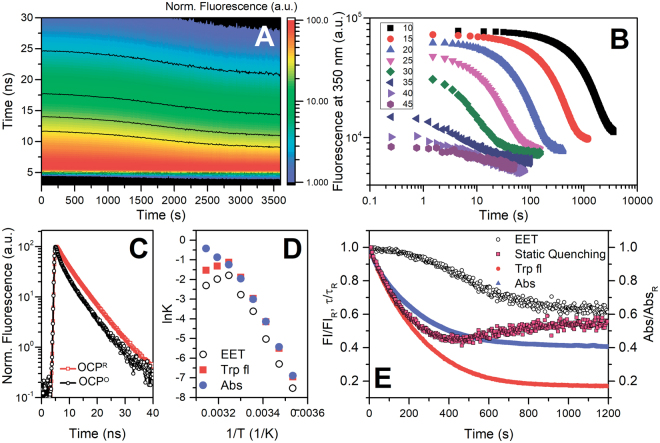
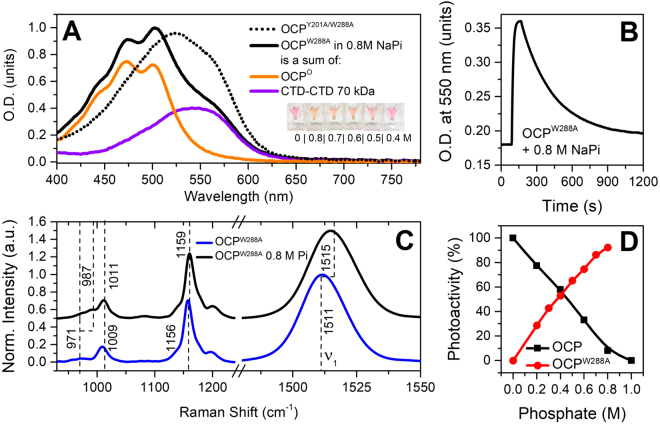
The 35-kDa Orange Carotenoid Protein (OCP) is responsible for photoprotection in cyanobacteria. It acts as a light intensity sensor and efficient quencher of phycobilisome excitation. Photoactivation triggers large-scale conformational rearrangements to convert OCP from the orange OCPO state to the red active signaling state, OCPR, as demonstrated by various structural methods. Such rearrangements imply a complete, yet reversible separation of structural domains and translocation of the carotenoid. Recently, dynamic crystallography of OCPO suggested the existence of photocycle intermediates with small-scale rearrangements that may trigger further transitions. In this study, we took advantage of single 7 ns laser pulses to study carotenoid absorption transients in OCP on the time-scale from 100 ns to 10 s, which allowed us to detect a red intermediate state preceding the red signaling state, OCPR. In addition, time-resolved fluorescence spectroscopy and the assignment of carotenoid-induced quenching of different tryptophan residues derived thereof revealed a novel orange intermediate state, which appears during the relaxation of photoactivated OCPR to OCPO. Our results show asynchronous changes between the carotenoid- and protein-associated kinetic components in a refined mechanistic model of the OCP photocycle, but also introduce new kinetic signatures for future studies of OCP photoactivity and photoprotection.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures


Similar articles
-
In vitro reconstitution of the cyanobacterial photoprotective mechanism mediated by the Orange Carotenoid Protein in Synechocystis PCC 6803.Plant Cell. 2011 Jul;23(7):2631-43. doi: 10.1105/tpc.111.086884. Epub 2011 Jul 15. Plant Cell. 2011. PMID: 21764991 Free PMC article.
-
Orange carotenoid protein burrows into the phycobilisome to provide photoprotection.Proc Natl Acad Sci U S A. 2016 Mar 22;113(12):E1655-62. doi: 10.1073/pnas.1523680113. Epub 2016 Mar 8. Proc Natl Acad Sci U S A. 2016. PMID: 26957606 Free PMC article.
-
Specificity of the cyanobacterial orange carotenoid protein: influences of orange carotenoid protein and phycobilisome structures.Plant Physiol. 2014 Feb;164(2):790-804. doi: 10.1104/pp.113.229997. Epub 2013 Dec 13. Plant Physiol. 2014. PMID: 24335507 Free PMC article.
-
The Orange Carotenoid Protein: a blue-green light photoactive protein.Photochem Photobiol Sci. 2013 Jul;12(7):1135-43. doi: 10.1039/c3pp25406b. Photochem Photobiol Sci. 2013. PMID: 23396391 Review.
-
Structure and functions of Orange Carotenoid Protein homologs in cyanobacteria.Curr Opin Plant Biol. 2017 Jun;37:1-9. doi: 10.1016/j.pbi.2017.03.010. Epub 2017 Apr 6. Curr Opin Plant Biol. 2017. PMID: 28391046 Review.
Cited by
-
Oligomerization processes limit photoactivation and recovery of the orange carotenoid protein.Biophys J. 2022 Aug 2;121(15):2849-2872. doi: 10.1016/j.bpj.2022.07.004. Epub 2022 Jul 6. Biophys J. 2022. PMID: 35794830 Free PMC article.
-
Probing of carotenoid-tryptophan hydrogen bonding dynamics in the single-tryptophan photoactive Orange Carotenoid Protein.Sci Rep. 2020 Jul 16;10(1):11729. doi: 10.1038/s41598-020-68463-8. Sci Rep. 2020. PMID: 32678150 Free PMC article.
-
Light-induced infrared difference spectroscopy on three different forms of orange carotenoid protein: focus on carotenoid vibrations.Photochem Photobiol Sci. 2023 Jun;22(6):1379-1391. doi: 10.1007/s43630-023-00384-7. Epub 2023 Feb 28. Photochem Photobiol Sci. 2023. PMID: 36853495
-
Comparative ultrafast spectroscopy and structural analysis of OCP1 and OCP2 from Tolypothrix.Biochim Biophys Acta Bioenerg. 2020 Feb 1;1861(2):148120. doi: 10.1016/j.bbabio.2019.148120. Epub 2019 Nov 14. Biochim Biophys Acta Bioenerg. 2020. PMID: 31734194 Free PMC article.
-
Changing Form and Function through Carotenoids and Synthetic Biology.Plant Physiol. 2019 Mar;179(3):830-843. doi: 10.1104/pp.18.01122. Epub 2018 Oct 25. Plant Physiol. 2019. PMID: 30361256 Free PMC article.
References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
