

Development and plasticity of meningeal lymphatic vessels
- PMID: 29141865
- PMCID: PMC5716035
- DOI: 10.1084/jem.20170391
Development and plasticity of meningeal lymphatic vessels
Abstract
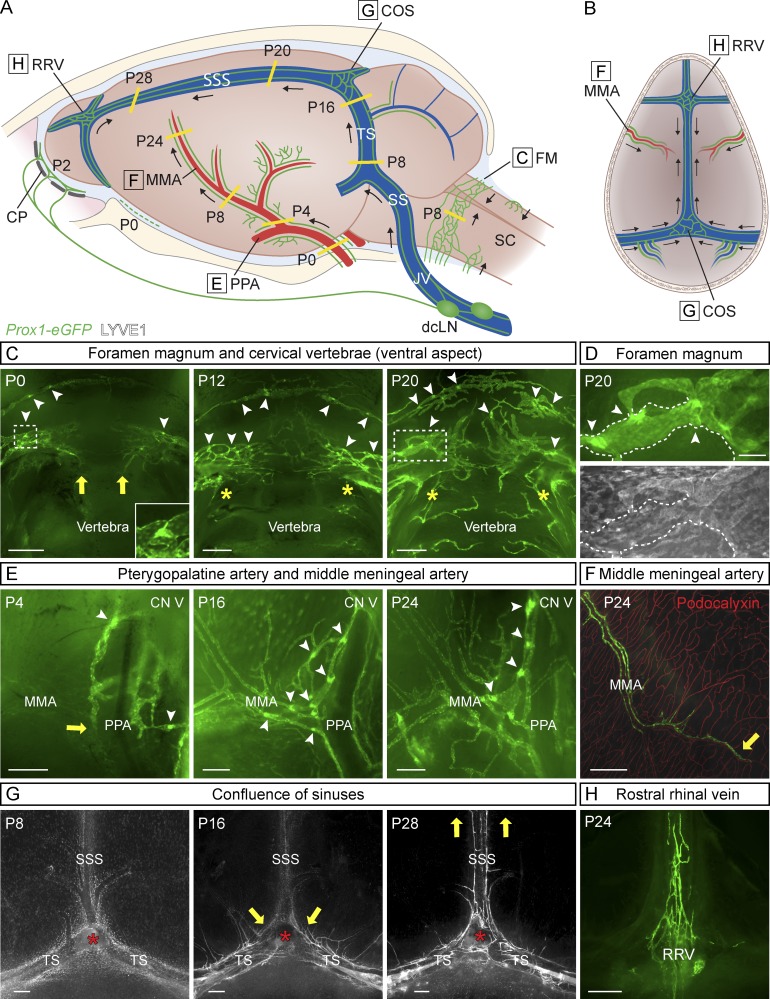
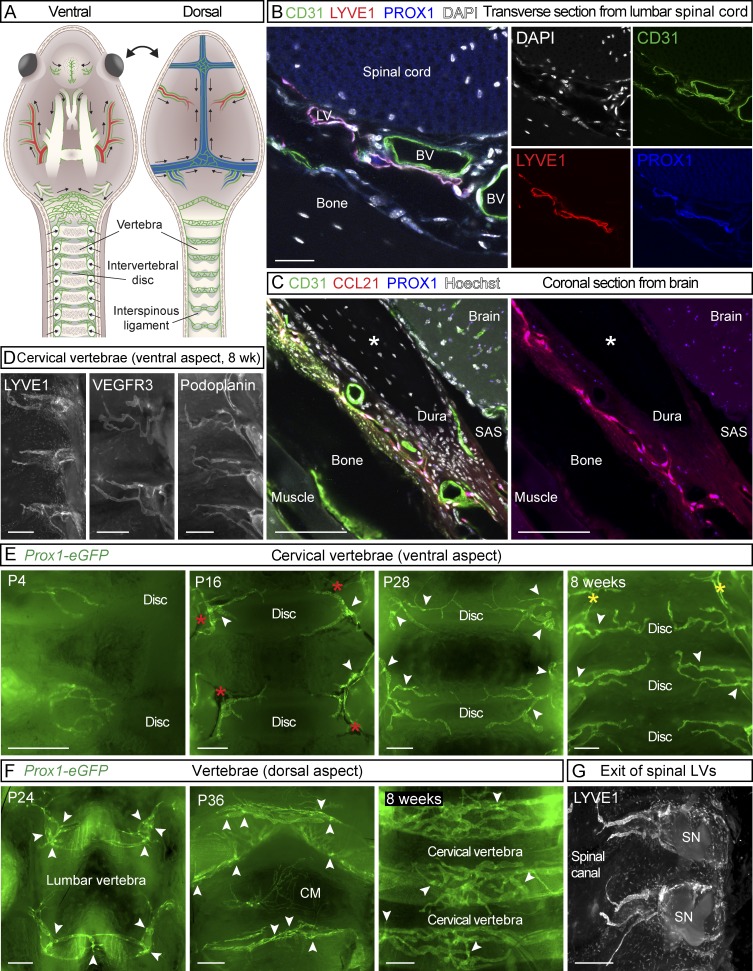
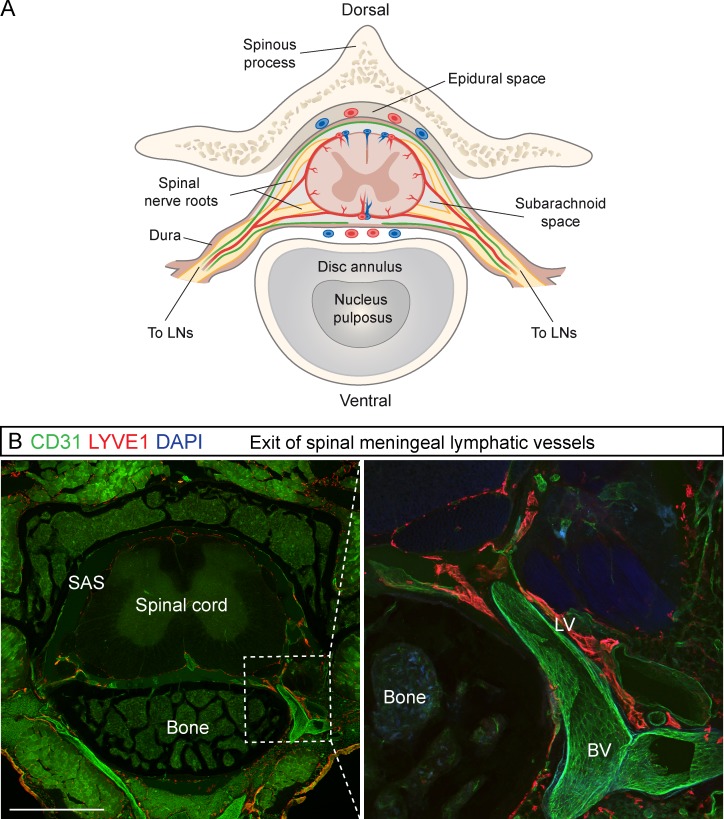
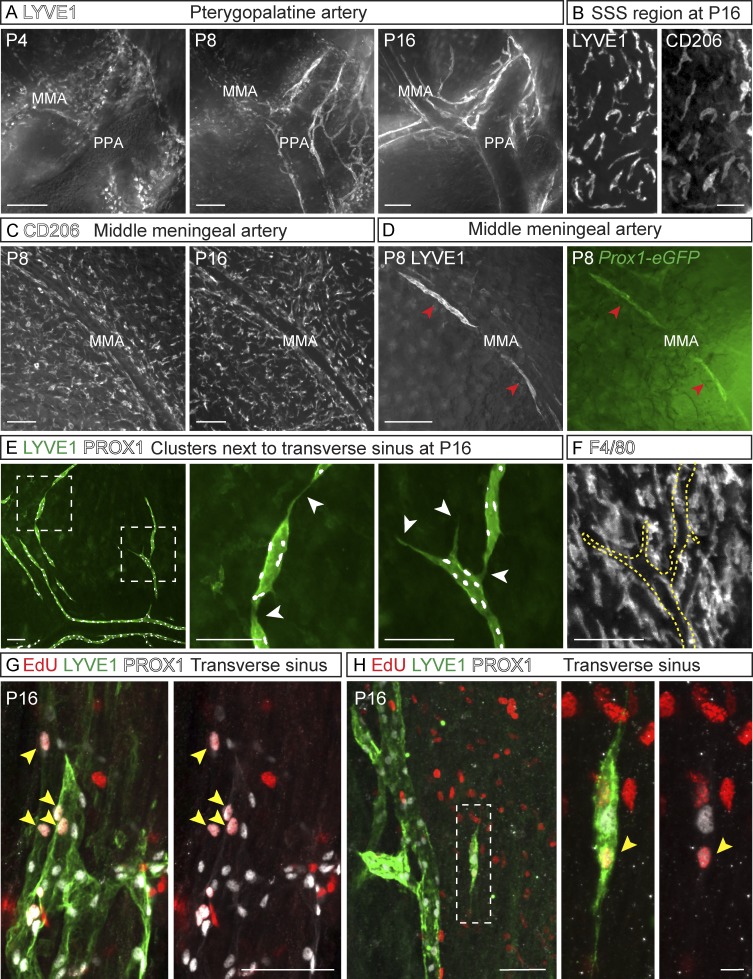
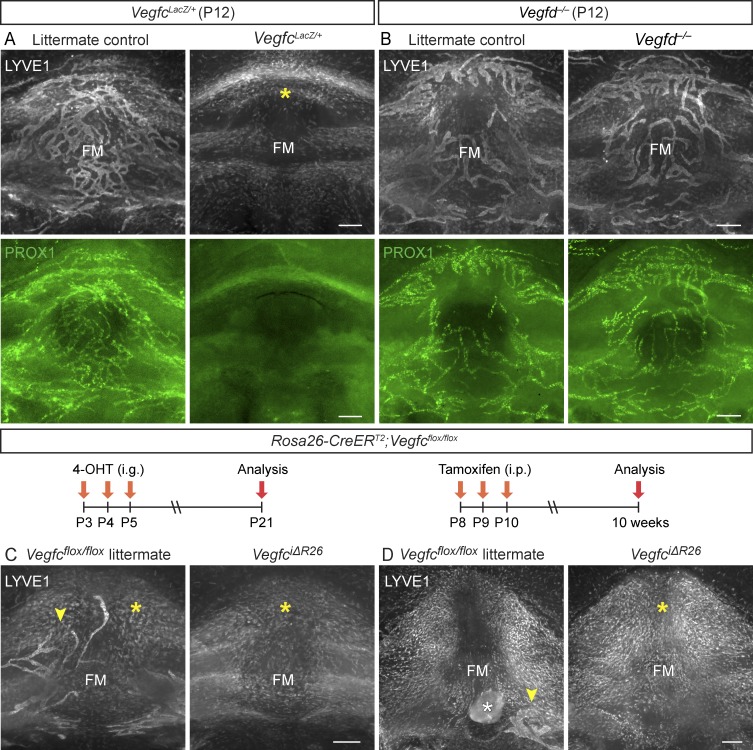
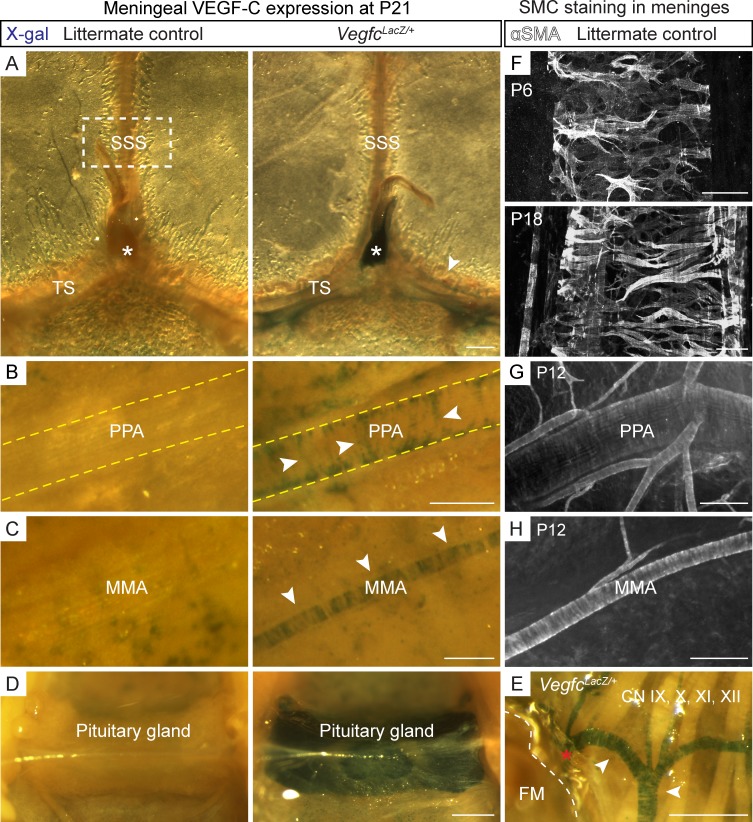
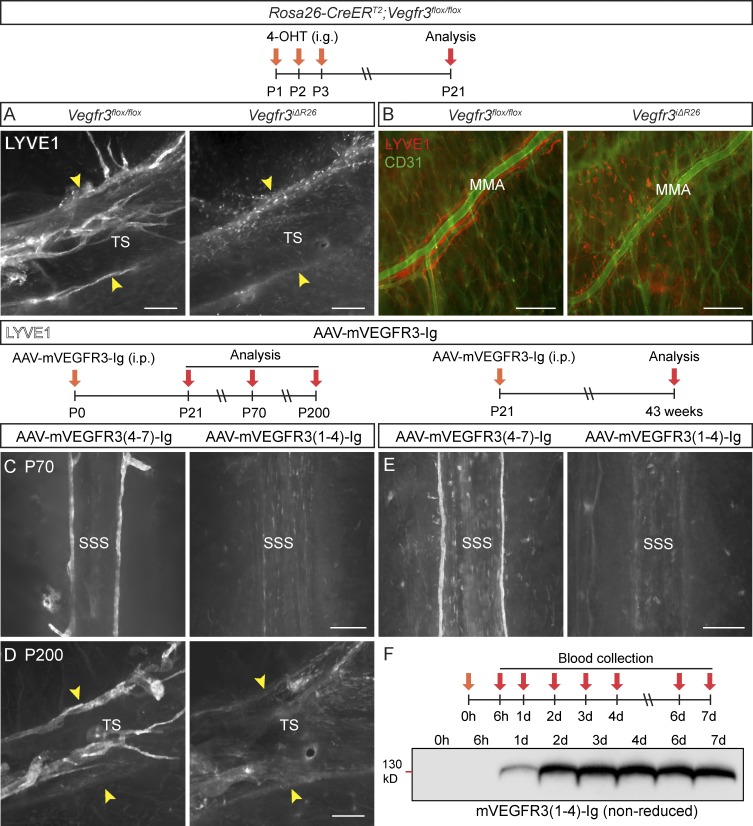
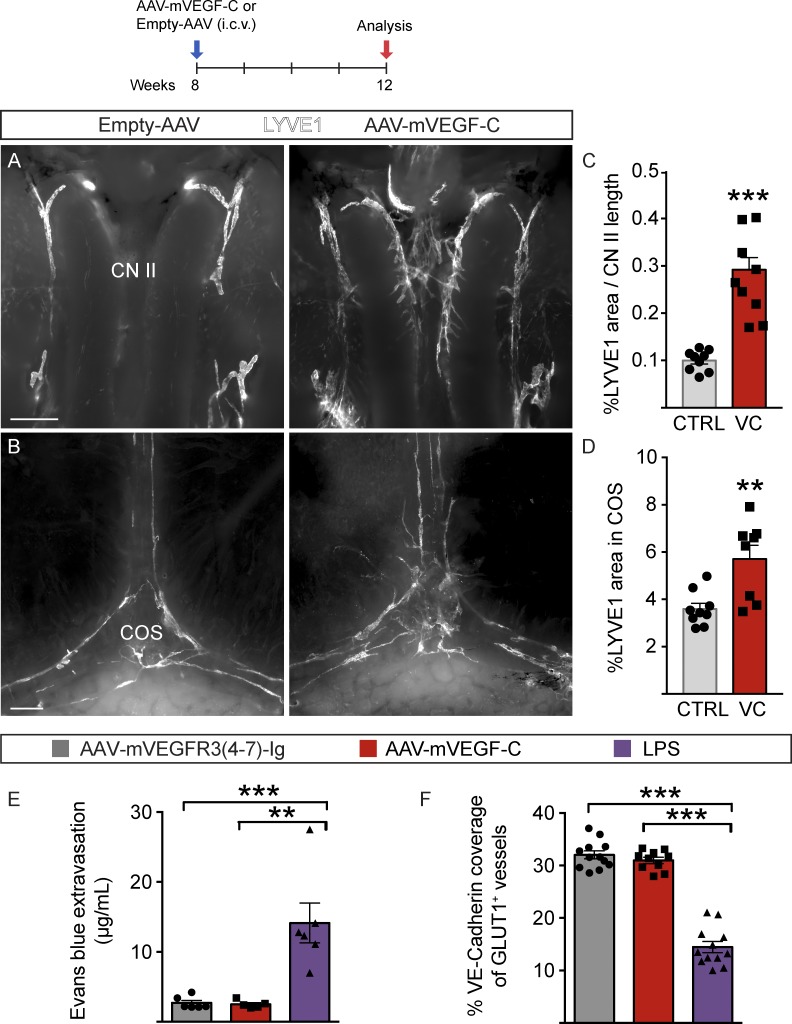
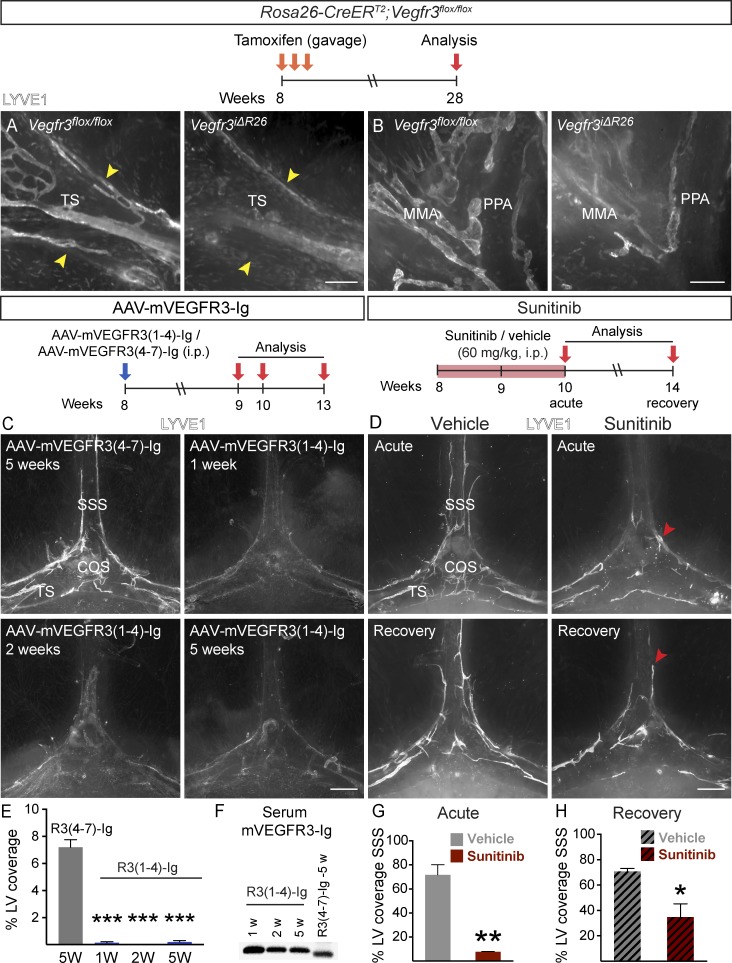
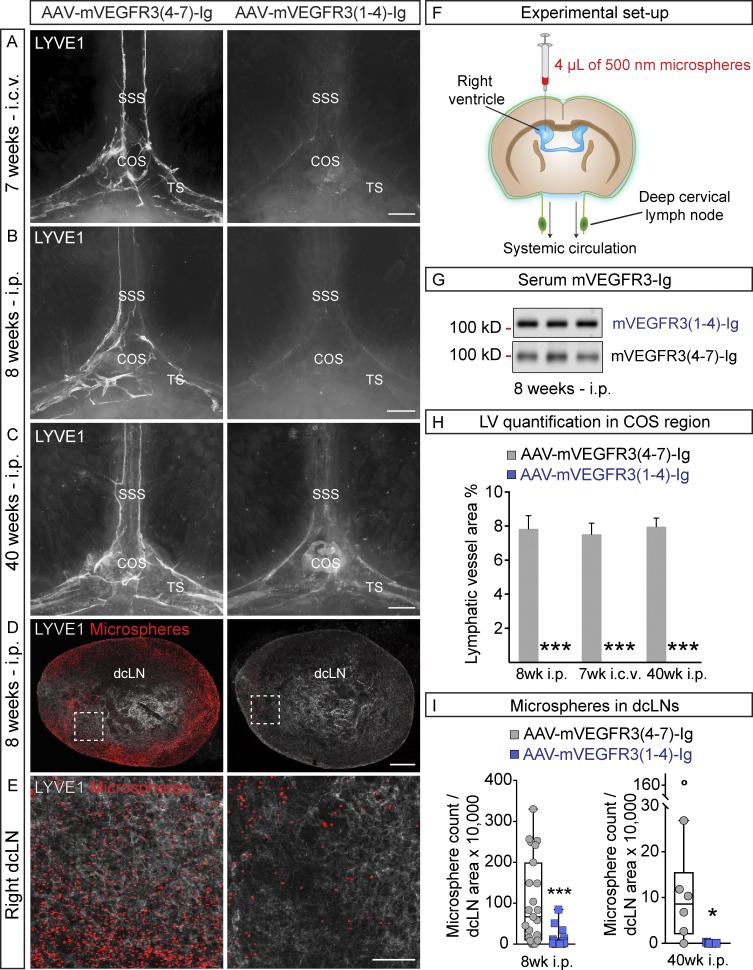
The recent discovery of meningeal lymphatic vessels (LVs) has raised interest in their possible involvement in neuropathological processes, yet little is known about their development or maintenance. We show here that meningeal LVs develop postnatally, appearing first around the foramina in the basal parts of the skull and spinal canal, sprouting along the blood vessels and cranial and spinal nerves to various parts of the meninges surrounding the central nervous system (CNS). VEGF-C, expressed mainly in vascular smooth muscle cells, and VEGFR3 in lymphatic endothelial cells were essential for their development, whereas VEGF-D deletion had no effect. Surprisingly, in adult mice, the LVs showed regression after VEGF-C or VEGFR3 deletion, administration of the tyrosine kinase inhibitor sunitinib, or expression of VEGF-C/D trap, which also compromised the lymphatic drainage function. Conversely, an excess of VEGF-C induced meningeal lymphangiogenesis. The plasticity and regenerative potential of meningeal LVs should allow manipulation of cerebrospinal fluid drainage and neuropathological processes in the CNS.
© 2017 Antila et al.
Figures
Comment in
-
The amazing brain drain.J Exp Med. 2017 Dec 4;214(12):3469-3470. doi: 10.1084/jem.20172031. Epub 2017 Nov 21. J Exp Med. 2017. PMID: 29162645 Free PMC article.
References
-
- Anisimov A., Alitalo A., Korpisalo P., Soronen J., Kaijalainen S., Leppänen V.M., Jeltsch M., Ylä-Herttuala S., and Alitalo K.. 2009. Activated forms of VEGF-C and VEGF-D provide improved vascular function in skeletal muscle. Circ. Res. 104:1302–1312. 10.1161/CIRCRESAHA.109.197830 - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
