

Effective Priming of Herpes Simplex Virus-Specific CD8+ T Cells In Vivo Does Not Require Infected Dendritic Cells
- PMID: 29142130
- PMCID: PMC5774880
- DOI: 10.1128/JVI.01508-17
Effective Priming of Herpes Simplex Virus-Specific CD8+ T Cells In Vivo Does Not Require Infected Dendritic Cells
Abstract
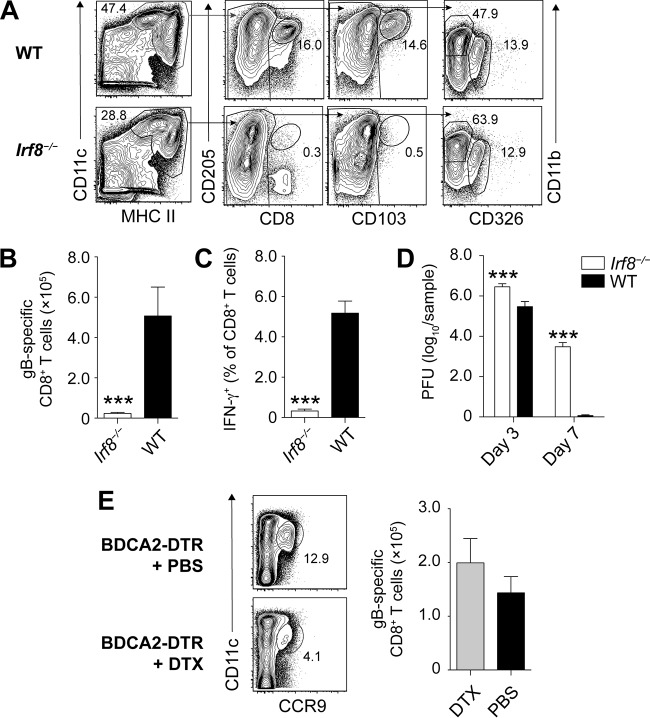
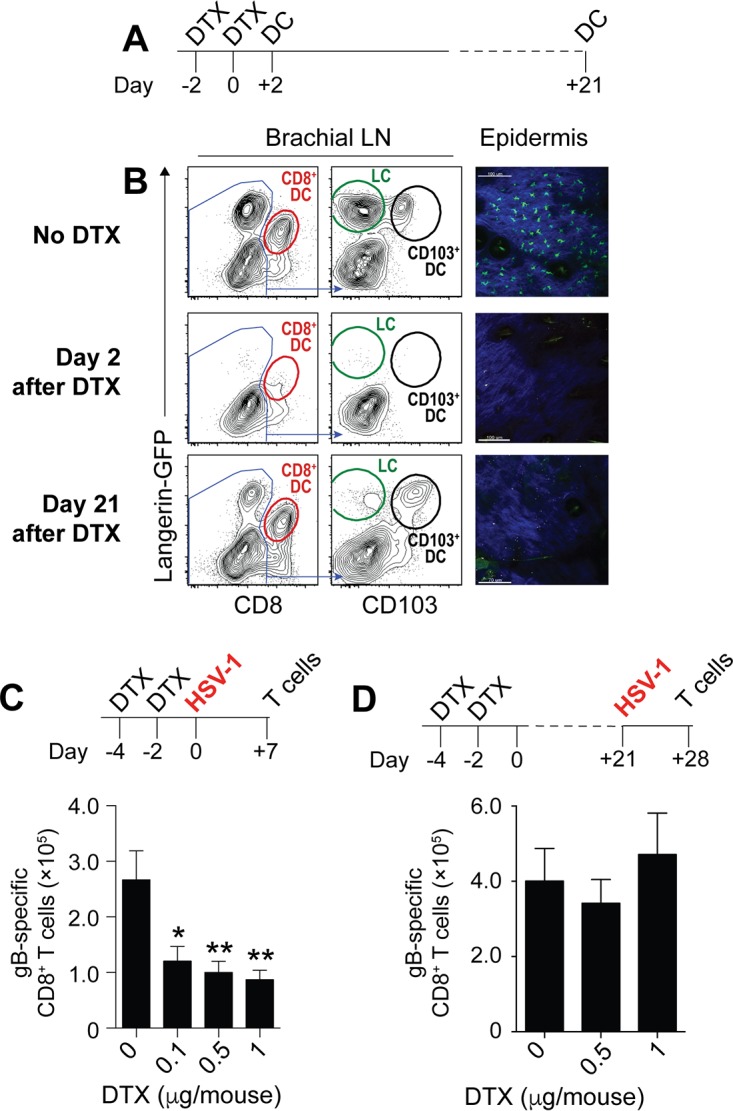
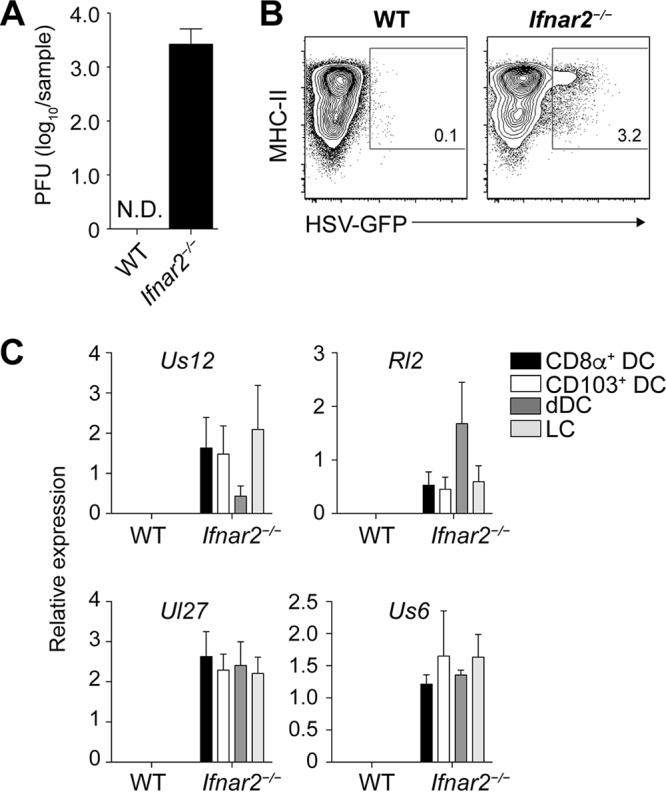
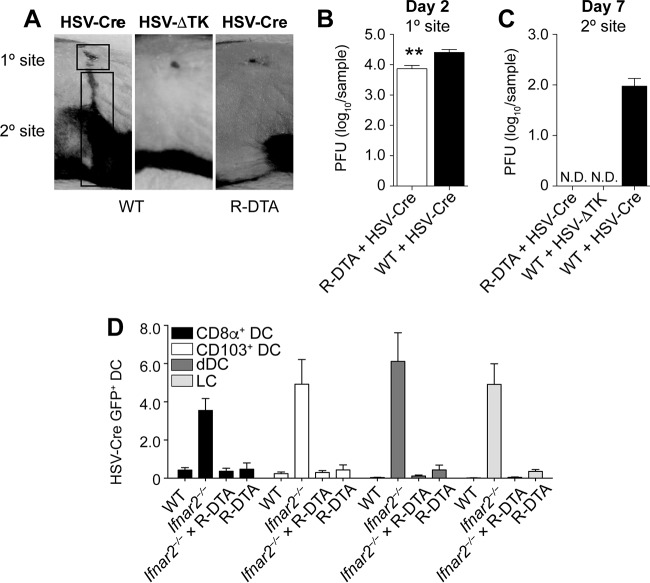
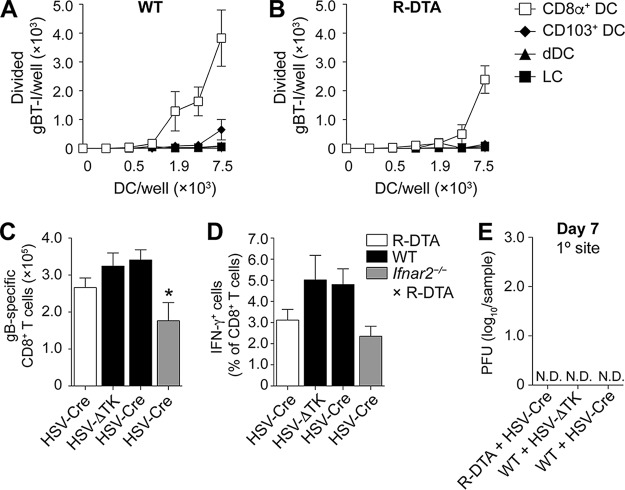
Resolution of virus infections depends on the priming of virus-specific CD8+ T cells by dendritic cells (DC). While this process requires major histocompatibility complex (MHC) class I-restricted antigen presentation by DC, the relative contribution to CD8+ T cell priming by infected DC is less clear. We have addressed this question in the context of a peripheral infection with herpes simplex virus 1 (HSV). Assessing the endogenous, polyclonal HSV-specific CD8+ T cell response, we found that effective in vivo T cell priming depended on the presence of DC subsets specialized in cross-presentation, while Langerhans cells and plasmacytoid DC were dispensable. Utilizing a novel mouse model that allows for the in vivo elimination of infected DC, we also demonstrated in vivo that this requirement for cross-presenting DC was not related to their infection but instead reflected their capacity to cross-present HSV-derived antigen. Taking the results together, this study shows that infected DC are not required for effective CD8+ T cell priming during a peripheral virus infection.IMPORTANCE The ability of some DC to present viral antigen to CD8+ T cells without being infected is thought to enable the host to induce killer T cells even when viruses evade or kill infected DC. However, direct experimental in vivo proof for this notion has remained elusive. The work described in this study characterizes the role that different DC play in the induction of virus-specific killer T cell responses and, critically, introduces a novel mouse model that allows for the selective elimination of infected DC in vivo Our finding that HSV-specific CD8+ T cells can be fully primed in the absence of DC infection shows that cross-presentation by DC is indeed sufficient for effective CD8+ T cell priming during a peripheral virus infection.
Keywords: CD8 T cells; HSV-1; cross-presentation; dendritic cells; direct presentation.
Copyright © 2018 American Society for Microbiology.
Figures
Similar articles
-
Contribution of direct and cross-presentation to CTL immunity against herpes simplex virus 1.J Immunol. 2009 Jan 1;182(1):283-92. doi: 10.4049/jimmunol.182.1.283. J Immunol. 2009. PMID: 19109159
-
Intrahepatic Cross-Presentation and Hepatocellular Antigen Presentation Play Distinct Roles in the Induction of Hepatitis B Virus-Specific CD8+ T Cell Responses.J Virol. 2018 Oct 12;92(21):e00920-18. doi: 10.1128/JVI.00920-18. Print 2018 Nov 1. J Virol. 2018. PMID: 30089700 Free PMC article.
-
Priming of CD8+ T cells against cytomegalovirus-encoded antigens is dominated by cross-presentation.J Immunol. 2013 Mar 15;190(6):2767-77. doi: 10.4049/jimmunol.1200966. Epub 2013 Feb 6. J Immunol. 2013. PMID: 23390296
-
Cross-presentation by dendritic cells.Nat Rev Immunol. 2012 Jul 13;12(8):557-69. doi: 10.1038/nri3254. Nat Rev Immunol. 2012. PMID: 22790179 Review.
-
TAP-ing into the cross-presentation secrets of dendritic cells.Curr Opin Immunol. 2023 Aug;83:102327. doi: 10.1016/j.coi.2023.102327. Epub 2023 Apr 26. Curr Opin Immunol. 2023. PMID: 37116384 Review.
Cited by
-
Different routes of MHC-I delivery to phagosomes and their consequences to CD8 T cell immunity.Semin Immunol. 2023 Mar;66:101713. doi: 10.1016/j.smim.2023.101713. Epub 2023 Jan 25. Semin Immunol. 2023. PMID: 36706521 Free PMC article. Review.
-
Initial TK-deficient HSV-1 infection in the lip alters contralateral lip challenge immune dynamics.Sci Rep. 2022 May 19;12(1):8489. doi: 10.1038/s41598-022-12597-4. Sci Rep. 2022. PMID: 35590057 Free PMC article.
-
The expression and function of HSV ICP47 and its promoter in mice.J Virol. 2023 Nov 30;97(11):e0110723. doi: 10.1128/jvi.01107-23. Epub 2023 Oct 30. J Virol. 2023. PMID: 37902400 Free PMC article.
-
Herpes simplex virus interference with immunity: Focus on dendritic cells.Virulence. 2021 Dec;12(1):2583-2607. doi: 10.1080/21505594.2021.1980990. Virulence. 2021. PMID: 34895058 Free PMC article. Review.
-
Mouse Mx1 Inhibits Herpes Simplex Virus Type 1 Genomic Replication and Late Gene Expression In Vitro and Prevents Lesion Formation in the Mouse Zosteriform Model.J Virol. 2022 Jun 22;96(12):e0041922. doi: 10.1128/jvi.00419-22. Epub 2022 May 31. J Virol. 2022. PMID: 35638820 Free PMC article.
References
-
- Bachem A, Guttler S, Hartung E, Ebstein F, Schaefer M, Tannert A, Salama A, Movassaghi K, Opitz C, Mages HW, Henn V, Kloetzel PM, Gurka S, Kroczek RA. 2010. Superior antigen cross-presentation and XCR1 expression define human CD11c+CD141+ cells as homologues of mouse CD8+ dendritic cells. J Exp Med 207:1273–1281. doi: 10.1084/jem.20100348. - DOI - PMC - PubMed
-
- Crozat K, Guiton R, Contreras V, Feuillet V, Dutertre CA, Ventre E, Vu Manh TP, Baranek T, Storset AK, Marvel J, Boudinot P, Hosmalin A, Schwartz-Cornil I, Dalod M. 2010. The XC chemokine receptor 1 is a conserved selective marker of mammalian cells homologous to mouse CD8alpha+ dendritic cells. J Exp Med 207:1283–1292. doi: 10.1084/jem.20100223. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
