

Cliques and cavities in the human connectome
- PMID: 29143250
- PMCID: PMC5769855
- DOI: 10.1007/s10827-017-0672-6
Cliques and cavities in the human connectome
Abstract
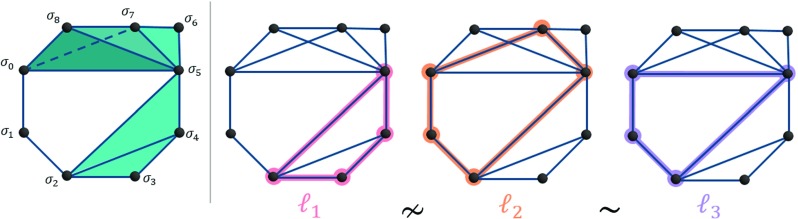
Encoding brain regions and their connections as a network of nodes and edges captures many of the possible paths along which information can be transmitted as humans process and perform complex behaviors. Because cognitive processes involve large, distributed networks of brain areas, principled examinations of multi-node routes within larger connection patterns can offer fundamental insights into the complexities of brain function. Here, we investigate both densely connected groups of nodes that could perform local computations as well as larger patterns of interactions that would allow for parallel processing. Finding such structures necessitates that we move from considering exclusively pairwise interactions to capturing higher order relations, concepts naturally expressed in the language of algebraic topology. These tools can be used to study mesoscale network structures that arise from the arrangement of densely connected substructures called cliques in otherwise sparsely connected brain networks. We detect cliques (all-to-all connected sets of brain regions) in the average structural connectomes of 8 healthy adults scanned in triplicate and discover the presence of more large cliques than expected in null networks constructed via wiring minimization, providing architecture through which brain network can perform rapid, local processing. We then locate topological cavities of different dimensions, around which information may flow in either diverging or converging patterns. These cavities exist consistently across subjects, differ from those observed in null model networks, and - importantly - link regions of early and late evolutionary origin in long loops, underscoring their unique role in controlling brain function. These results offer a first demonstration that techniques from algebraic topology offer a novel perspective on structural connectomics, highlighting loop-like paths as crucial features in the human brain's structural architecture.
Keywords: Applied topology; Network neuroscience; Persistent homology.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures


























References
-
- Bassett DS, Greenfield DL, Meyer-Lindenberg A, Weinberger DR, Moore SW, Bullmore ET. Efficient physical embedding of topologically complex information processing networks in brains and computer circuits. PLoS Computational Biology. 2010;6(4):1000748. doi: 10.1371/journal.pcbi.1000748. - DOI - PMC - PubMed
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
