

Innately activated TLR4 signal in the nucleus accumbens is sustained by CRF amplification loop and regulates impulsivity
- PMID: 29146239
- PMCID: PMC5857415
- DOI: 10.1016/j.bbi.2017.11.008
Innately activated TLR4 signal in the nucleus accumbens is sustained by CRF amplification loop and regulates impulsivity
Abstract
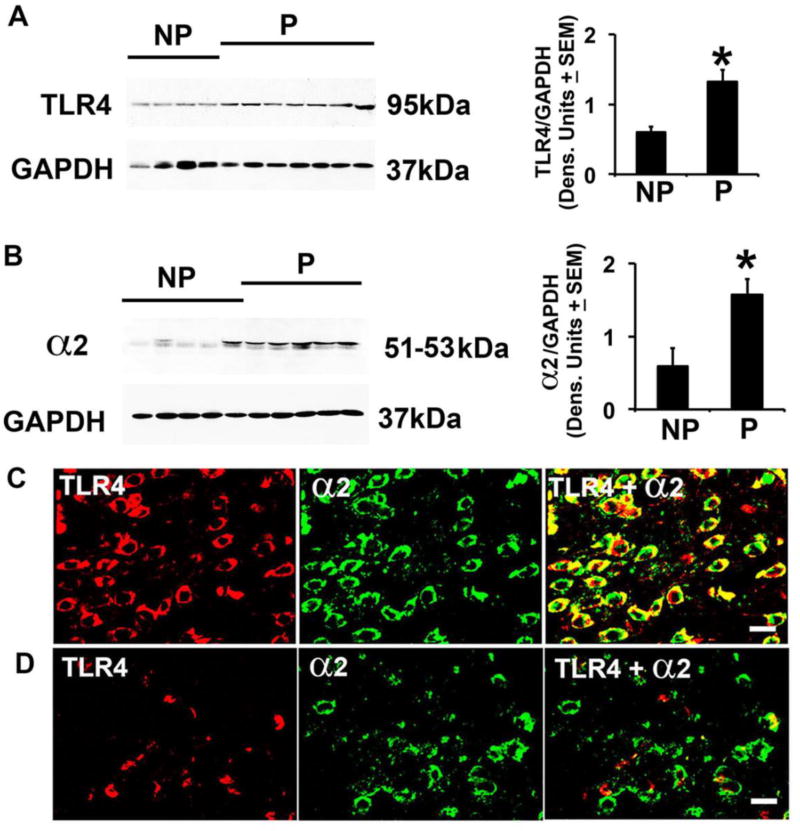
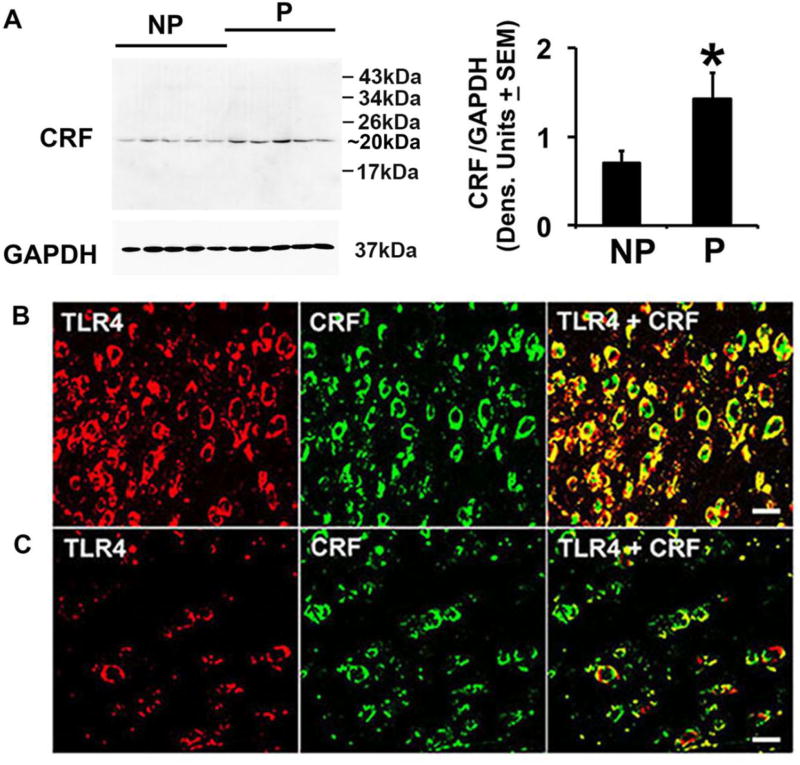
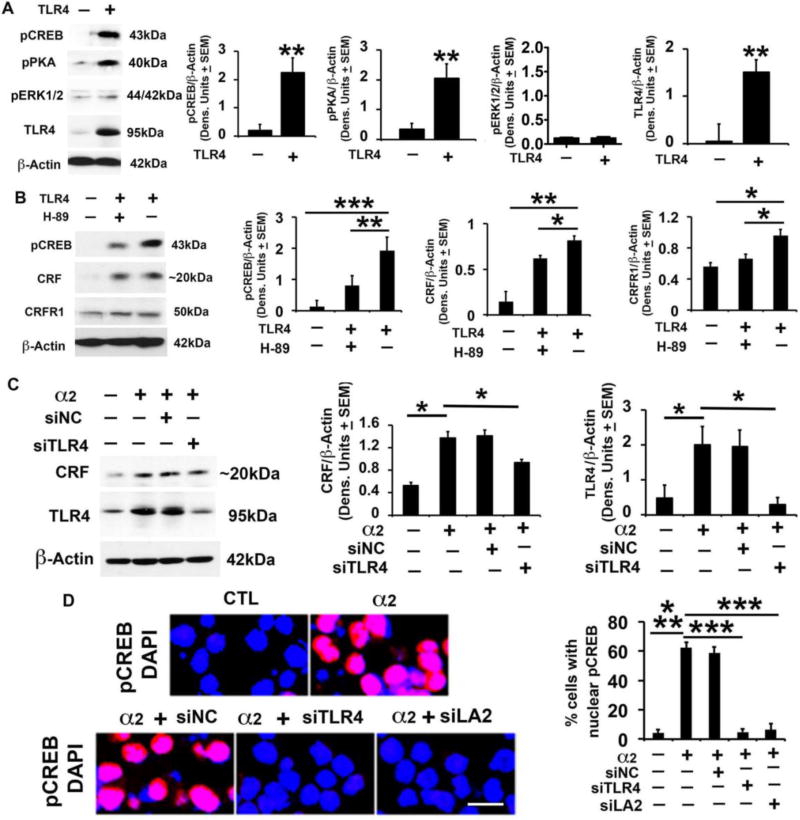
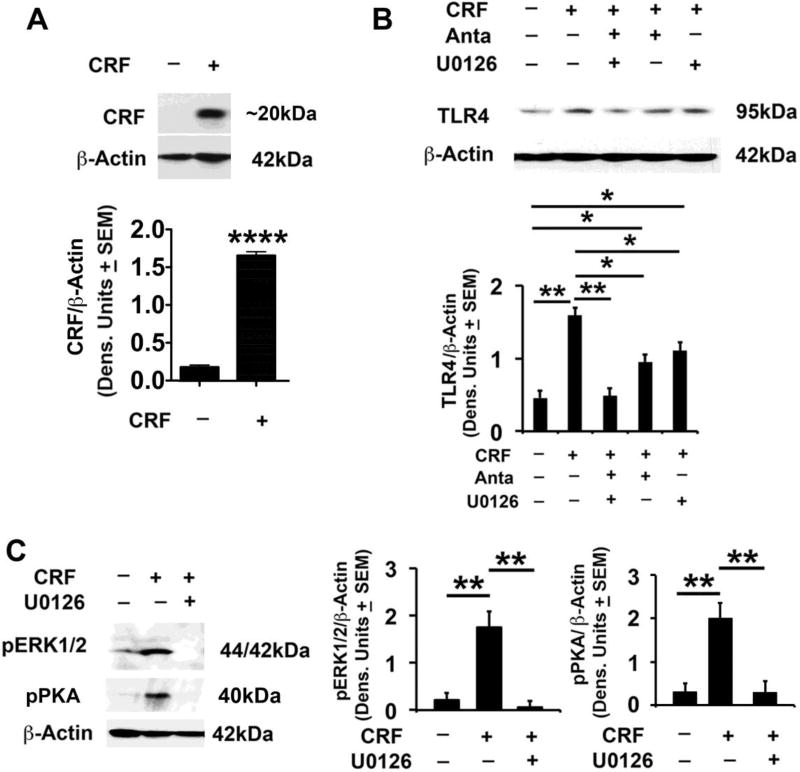
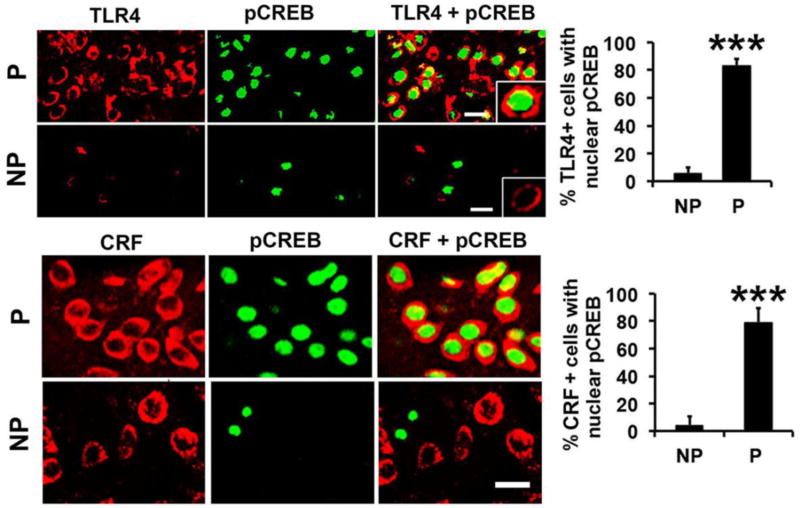
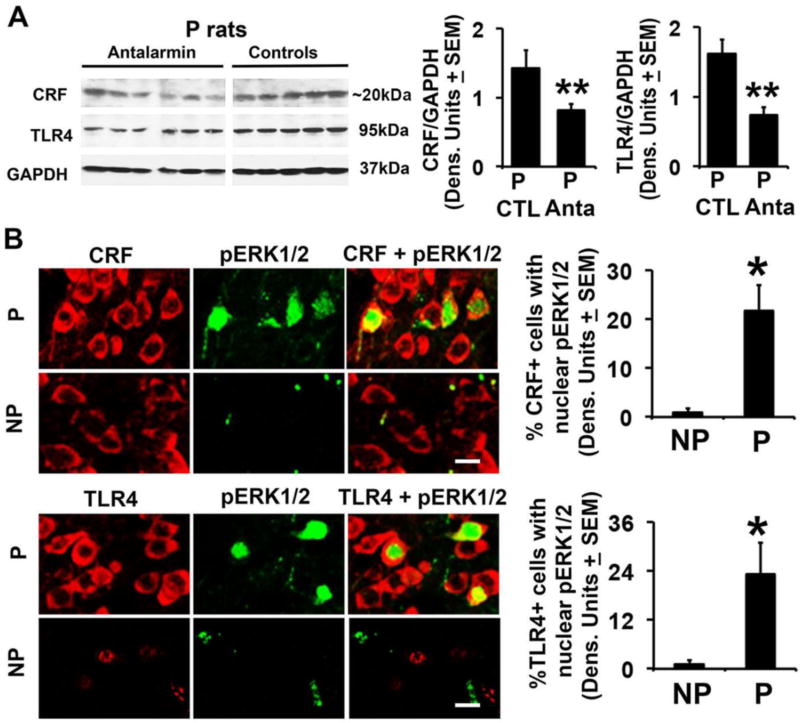
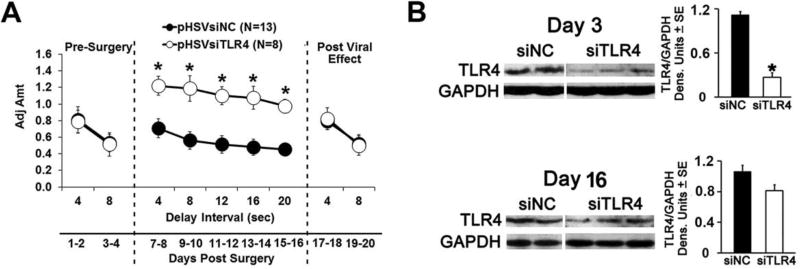
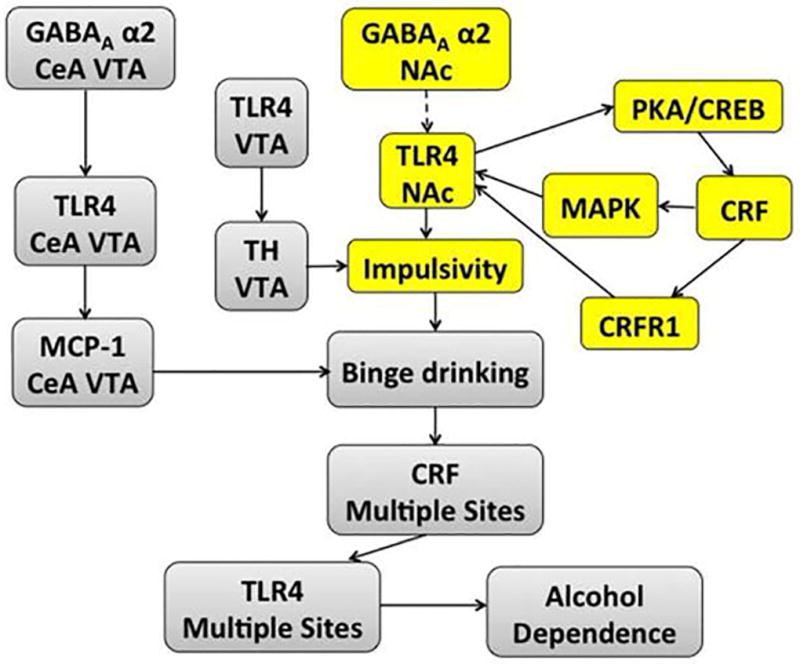
Cognitive impulsivity is a heritable trait believed to represent the behavior that defines the volition to initiate alcohol drinking. We have previously shown that a neuronal Toll-like receptor 4 (TLR4) signal located in the central amygdala (CeA) and ventral tegmental area (VTA) controls the initiation of binge drinking in alcohol-preferring P rats, and TLR4 expression is upregulated by alcohol-induced corticotropin-releasing factor (CRF) at these sites. However, the function of the TLR4 signal in the nucleus accumbens shell (NAc-shell), a site implicated in the control of reward, drug-seeking behavior and impulsivity and the contribution of other signal-associated genes, are still poorly understood. Here we report that P rats have an innately activated TLR4 signal in NAc-shell neurons that co-express the α2 GABAA receptor subunit and CRF prior to alcohol exposure. This signal is not present in non-alcohol drinking NP rats. The TLR4 signal is sustained by a CRF amplification loop, which includes TLR4-mediated CRF upregulation through PKA/CREB activation and CRF-mediated TLR4 upregulation through the CRF type 1 receptor (CRFR1) and the MAPK/ERK pathway. NAc-shell Infusion of a neurotropic, non-replicating herpes simplex virus vector for TLR4-specific small interfering RNA (pHSVsiTLR4) inhibits TLR4 expression and cognitive impulsivity, implicating the CRF-amplified TLR4 signal in impulsivity regulation.
Keywords: Activated TLR4 signal; CRF; GABA(A) α2; HSV siRNA vectors; Impulsivity; PKA/CREB.
Copyright © 2017 Elsevier Inc. All rights reserved.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Ahmed BY, Husnain O, Stafford R, Howard M, Gujar AS, Moradiya V, Patel KK, Sihotra S. Hyperphosphorylation of CREB in human dopaminergic neurons: a kinetic study of cellular distribution of total CREB and phospho-CREB following oxidative stress. Neuroreport. 2013;24:757–762. - PubMed
-
- Alfonso-Loeches S, Urena-Peralta J, Morillo-Bargues MJ, Gomez-Pinedo U, Guerri C. Ethanol-Induced TLR4/NLRP3 Neuroinflammatory Response in Microglial Cells Promotes Leukocyte Infiltration Across the BBB. Neurochem Res. 2016;41:193–209. - PubMed
-
- Asher O, Cunningham TD, Yao L, Gordon AS, Diamond I. Ethanol stimulates cAMP-responsive element (CRE)-mediated transcription via CRE-binding protein and cAMP-dependent protein kinase. J Pharmacol Exp Ther. 2002;301:66–70. - PubMed
-
- Aurelian L. Herpes Simplex Viruses: General Features. In: Mahy BWJ, van Regenmortel MHV, editors. Encyclopedia of Virology. 3. Elsevier, ltd; 2014. pp. 383–397.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
