

The structural basis of flagellin detection by NAIP5: A strategy to limit pathogen immune evasion
- PMID: 29146805
- PMCID: PMC5842810
- DOI: 10.1126/science.aao1140
The structural basis of flagellin detection by NAIP5: A strategy to limit pathogen immune evasion
Abstract
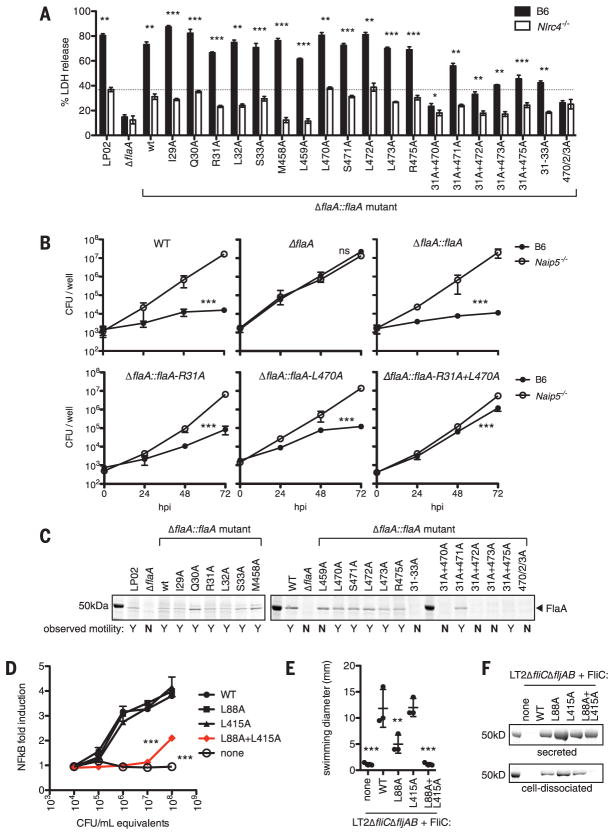
Robust innate immune detection of rapidly evolving pathogens is critical for host defense. Nucleotide-binding domain leucine-rich repeat (NLR) proteins function as cytosolic innate immune sensors in plants and animals. However, the structural basis for ligand-induced NLR activation has so far remained unknown. NAIP5 (NLR family, apoptosis inhibitory protein 5) binds the bacterial protein flagellin and assembles with NLRC4 to form a multiprotein complex called an inflammasome. Here we report the cryo-electron microscopy structure of the assembled ~1.4-megadalton flagellin-NAIP5-NLRC4 inflammasome, revealing how a ligand activates an NLR. Six distinct NAIP5 domains contact multiple conserved regions of flagellin, prying NAIP5 into an open and active conformation. We show that innate immune recognition of multiple ligand surfaces is a generalizable strategy that limits pathogen evolution and immune escape.
Copyright © 2017, American Association for the Advancement of Science.
Figures
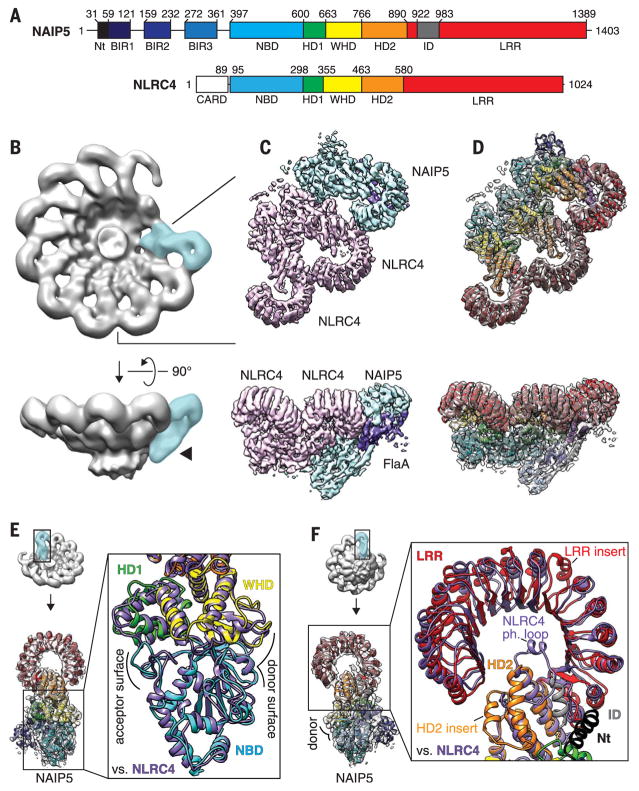
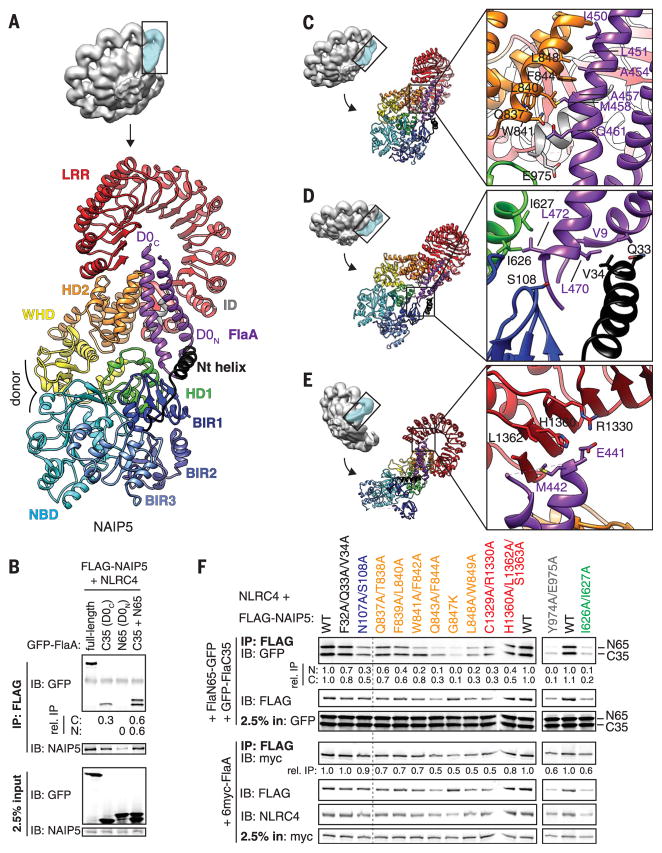
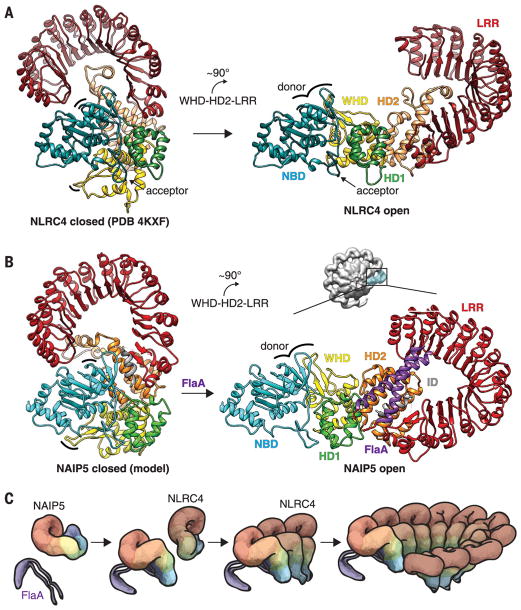
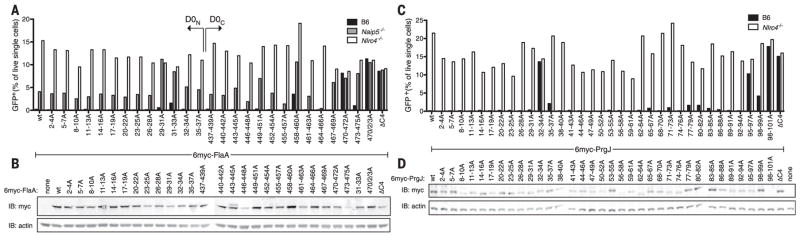
Comment in
-
Bad germs are trapped.Cell Res. 2018 Feb;28(2):141-142. doi: 10.1038/cr.2018.5. Epub 2018 Jan 9. Cell Res. 2018. PMID: 29313529 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
