

Postcatalytic spliceosome structure reveals mechanism of 3'-splice site selection
- PMID: 29146871
- PMCID: PMC5808836
- DOI: 10.1126/science.aar3729
Postcatalytic spliceosome structure reveals mechanism of 3'-splice site selection
Abstract
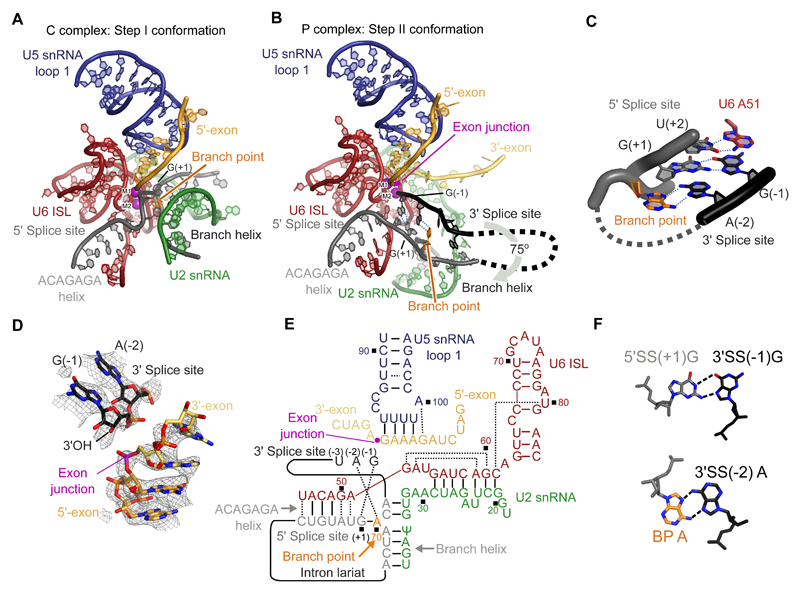
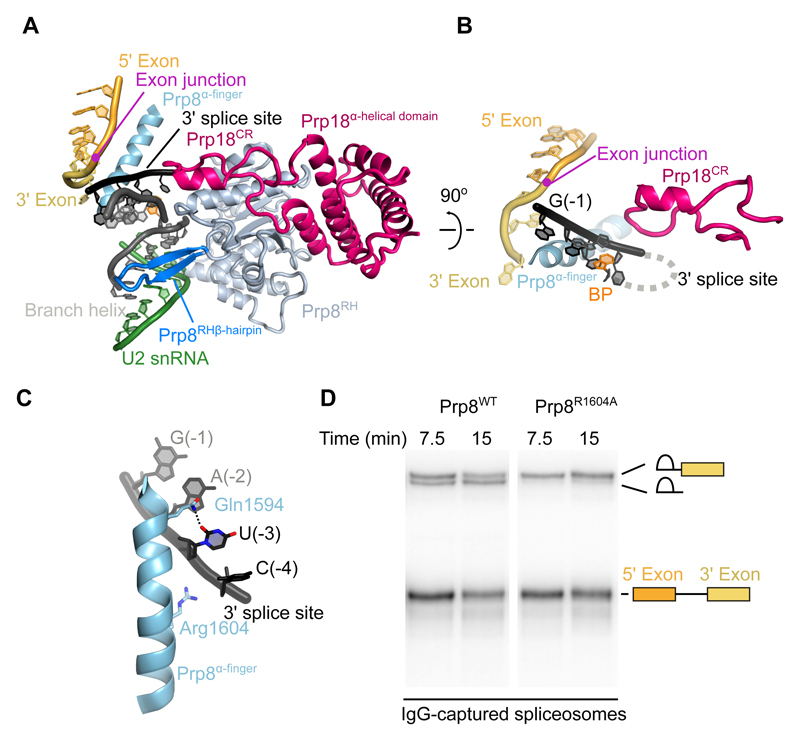
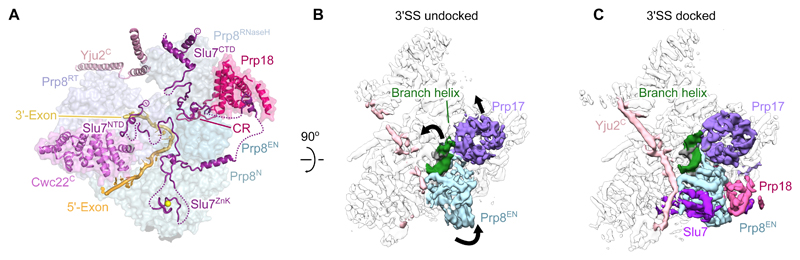
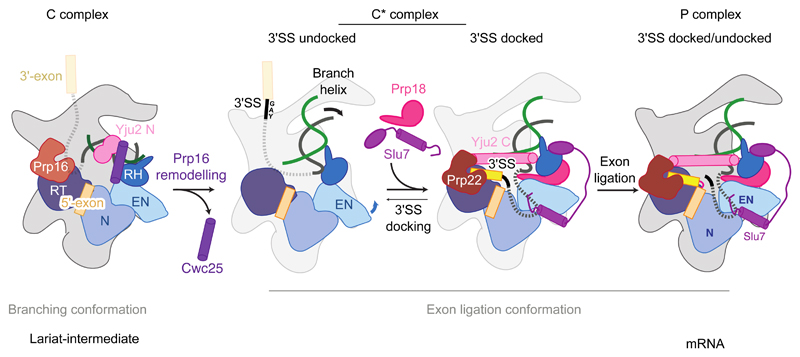
Introns are removed from eukaryotic messenger RNA precursors by the spliceosome in two transesterification reactions-branching and exon ligation. The mechanism of 3'-splice site recognition during exon ligation has remained unclear. Here we present the 3.7-angstrom cryo-electron microscopy structure of the yeast P-complex spliceosome immediately after exon ligation. The 3'-splice site AG dinucleotide is recognized through non-Watson-Crick pairing with the 5' splice site and the branch-point adenosine. After the branching reaction, protein factors work together to remodel the spliceosome and stabilize a conformation competent for 3'-splice site docking, thereby promoting exon ligation. The structure accounts for the strict conservation of the GU and AG dinucleotides at the 5' and 3' ends of introns and provides insight into the catalytic mechanism of exon ligation.
Copyright © 2017 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works.
Figures

Similar articles
-
Cryo-EM structure of the spliceosome immediately after branching.Nature. 2016 Sep 8;537(7619):197-201. doi: 10.1038/nature19316. Epub 2016 Jul 26. Nature. 2016. PMID: 27459055 Free PMC article.
-
Structure of a spliceosome remodelled for exon ligation.Nature. 2017 Feb 16;542(7641):377-380. doi: 10.1038/nature21078. Epub 2017 Jan 11. Nature. 2017. PMID: 28076345 Free PMC article.
-
Structure of the yeast spliceosomal postcatalytic P complex.Science. 2017 Dec 8;358(6368):1278-1283. doi: 10.1126/science.aar3462. Epub 2017 Nov 16. Science. 2017. PMID: 29146870 Free PMC article.
-
How Is Precursor Messenger RNA Spliced by the Spliceosome?Annu Rev Biochem. 2020 Jun 20;89:333-358. doi: 10.1146/annurev-biochem-013118-111024. Epub 2019 Dec 9. Annu Rev Biochem. 2020. PMID: 31815536 Review.
-
Molecular choreography of pre-mRNA splicing by the spliceosome.Curr Opin Struct Biol. 2019 Dec;59:124-133. doi: 10.1016/j.sbi.2019.07.010. Epub 2019 Aug 30. Curr Opin Struct Biol. 2019. PMID: 31476650 Review.
Cited by
-
Structural basis for conformational equilibrium of the catalytic spliceosome.Mol Cell. 2021 Apr 1;81(7):1439-1452.e9. doi: 10.1016/j.molcel.2021.02.021. Epub 2021 Mar 10. Mol Cell. 2021. PMID: 33705709 Free PMC article.
-
Termination of pre-mRNA splicing requires that the ATPase and RNA unwindase Prp43p acts on the catalytic snRNA U6.Genes Dev. 2019 Nov 1;33(21-22):1555-1574. doi: 10.1101/gad.328294.119. Epub 2019 Sep 26. Genes Dev. 2019. PMID: 31558568 Free PMC article.
-
Ranking noncanonical 5' splice site usage by genome-wide RNA-seq analysis and splicing reporter assays.Genome Res. 2018 Dec;28(12):1826-1840. doi: 10.1101/gr.235861.118. Epub 2018 Oct 24. Genome Res. 2018. PMID: 30355602 Free PMC article.
-
Animal, Fungi, and Plant Genome Sequences Harbor Different Non-Canonical Splice Sites.Cells. 2020 Feb 18;9(2):458. doi: 10.3390/cells9020458. Cells. 2020. PMID: 32085510 Free PMC article.
-
Multi-step recognition of potential 5' splice sites by the Saccharomyces cerevisiae U1 snRNP.Elife. 2022 Aug 12;11:e70534. doi: 10.7554/eLife.70534. Elife. 2022. PMID: 35959885 Free PMC article.
References
-
- Wahl MC, Will CL, Lührmann R. The Spliceosome: Design Principles of a Dynamic RNP Machine. Cell. 2009;136:701–718. - PubMed
-
- Patel AA, Steitz JA. Splicing double: insights from the second spliceosome. Nat Rev Mol Cell Biol. 2003;4:960–970. - PubMed
-
- Schwer B, Guthrie C. PRP16 is an RNA-dependent ATPase that interacts transiently with the spliceosome. Nature. 1991;349:494–499. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
