

Holistic bioengineering: rewiring central metabolism for enhanced bioproduction
- PMID: 29146872
- PMCID: PMC5688466
- DOI: 10.1042/BCJ20170377
Holistic bioengineering: rewiring central metabolism for enhanced bioproduction
Abstract
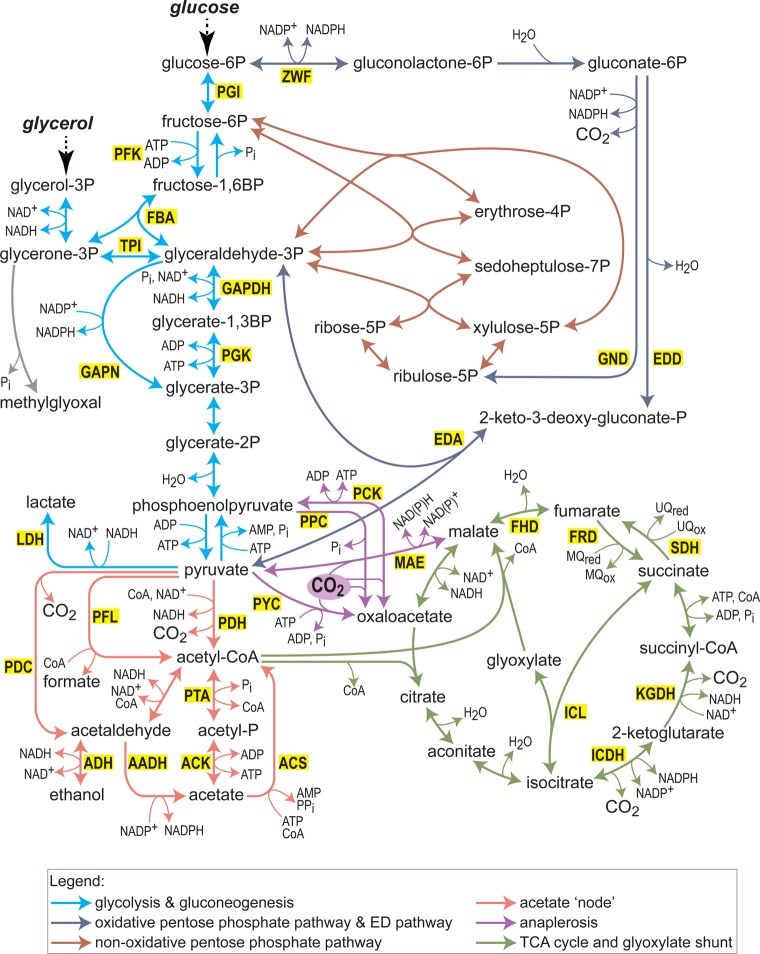
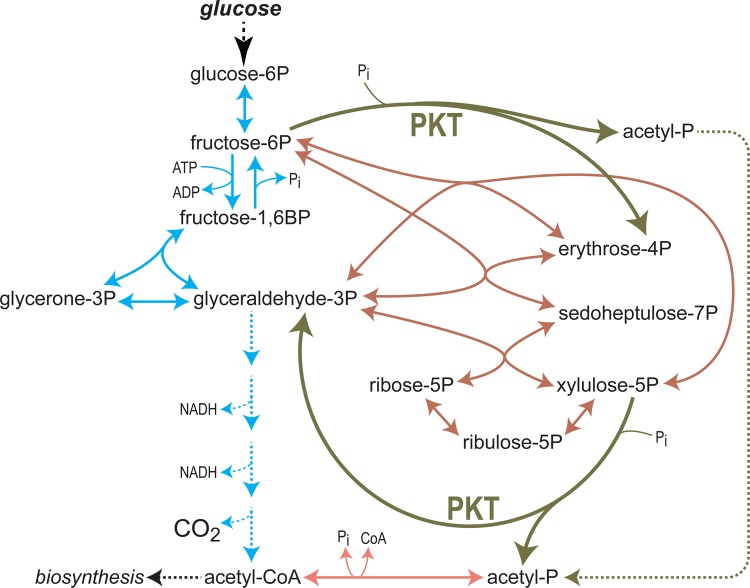
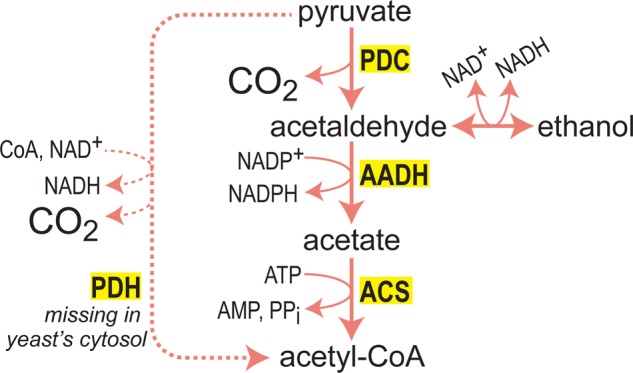
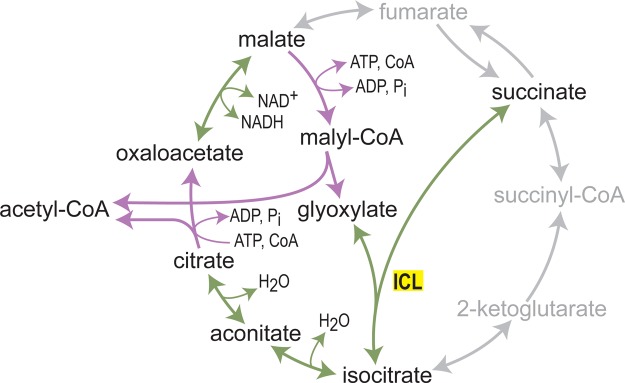
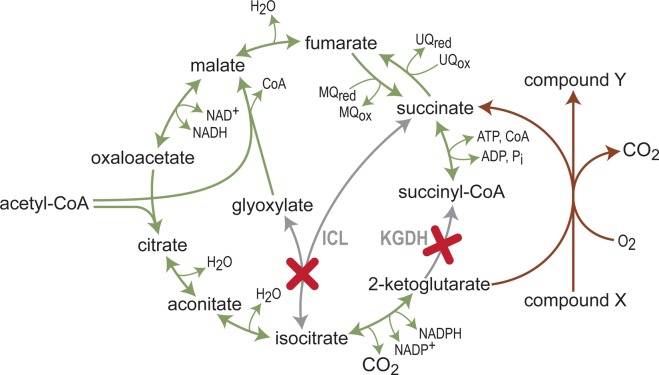
What does it take to convert a living organism into a truly productive biofactory? Apart from optimizing biosynthesis pathways as standalone units, a successful bioengineering approach must bend the endogenous metabolic network of the host, and especially its central metabolism, to support the bioproduction process. In practice, this usually involves three complementary strategies which include tuning-down or abolishing competing metabolic pathways, increasing the availability of precursors of the desired biosynthesis pathway, and ensuring high availability of energetic resources such as ATP and NADPH. In this review, we explore these strategies, focusing on key metabolic pathways and processes, such as glycolysis, anaplerosis, the TCA (tricarboxylic acid) cycle, and NADPH production. We show that only a holistic approach for bioengineering - considering the metabolic network of the host organism as a whole, rather than focusing on the production pathway alone - can truly mold microorganisms into efficient biofactories.
Keywords: NADPH production; TCA cycle; anaplerosis; central metabolism; glycolysis; metabolic engineering.
© 2017 The Author(s).
Conflict of interest statement
The Authors declare that there are no competing interests associated with the manuscript.
Figures

References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources