

Discovering the 3' UTR-mediated regulation of alpha-synuclein
- PMID: 29149290
- PMCID: PMC5728410
- DOI: 10.1093/nar/gkx1048
Discovering the 3' UTR-mediated regulation of alpha-synuclein
Abstract
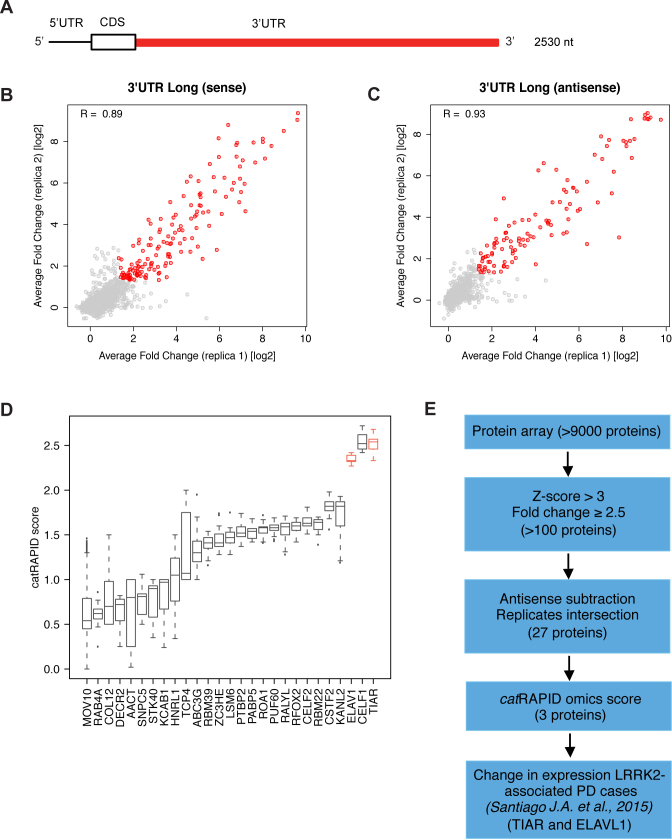
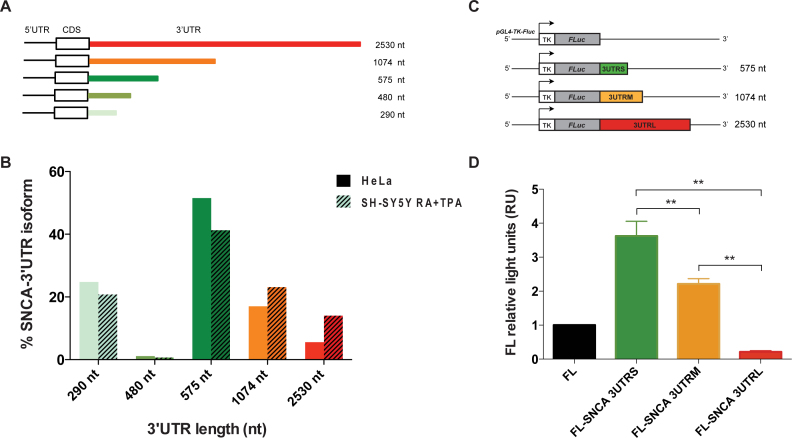
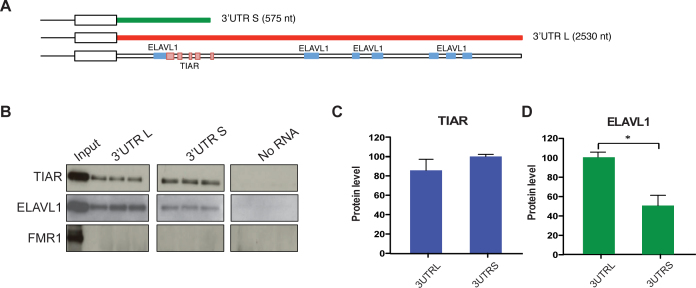
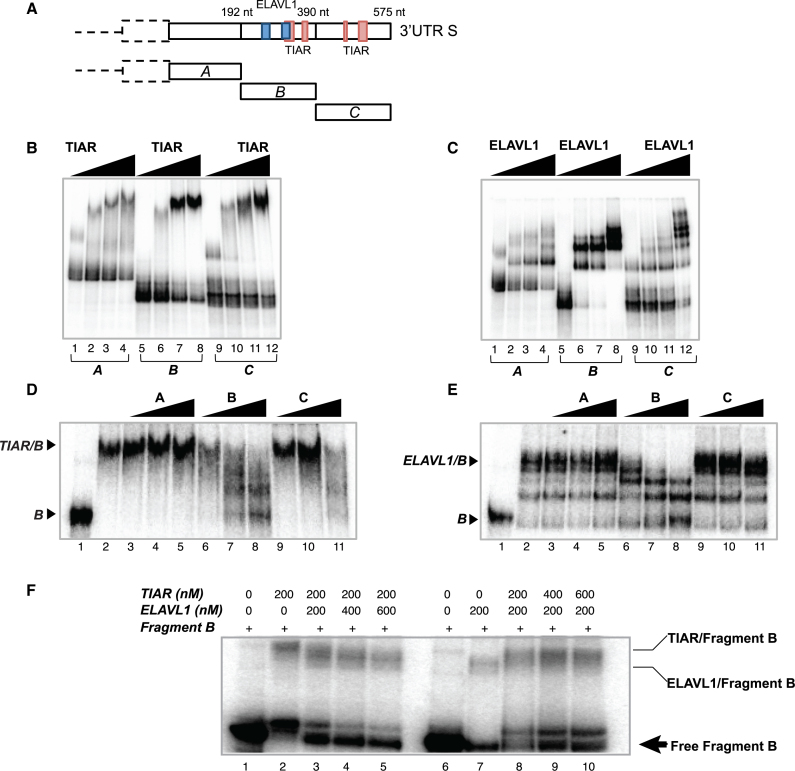
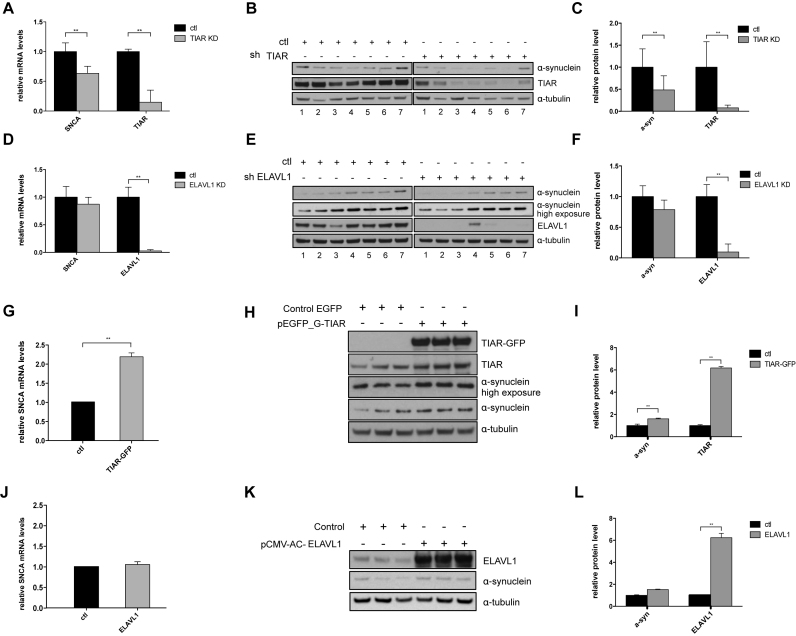
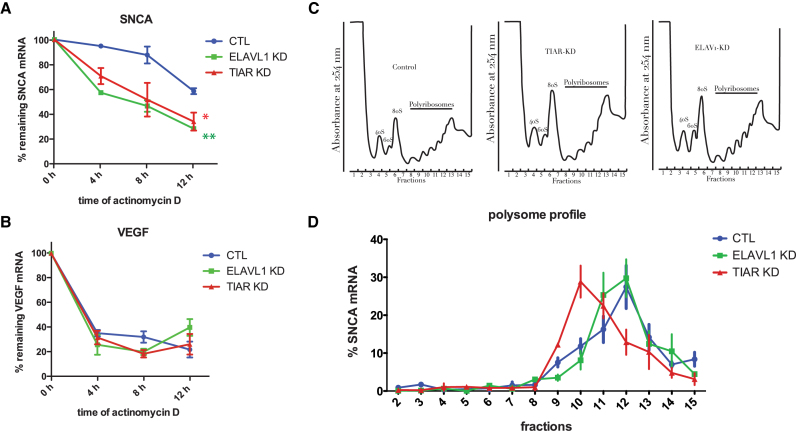
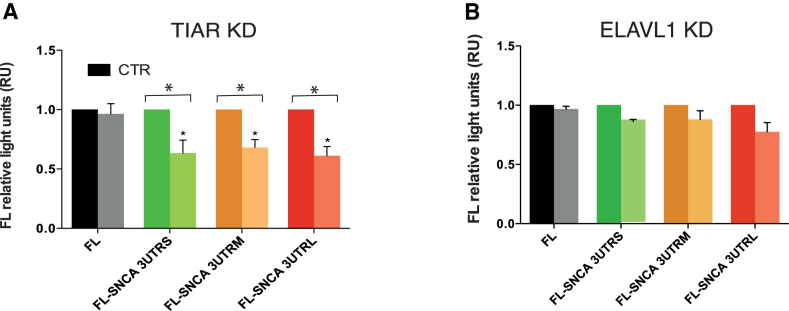
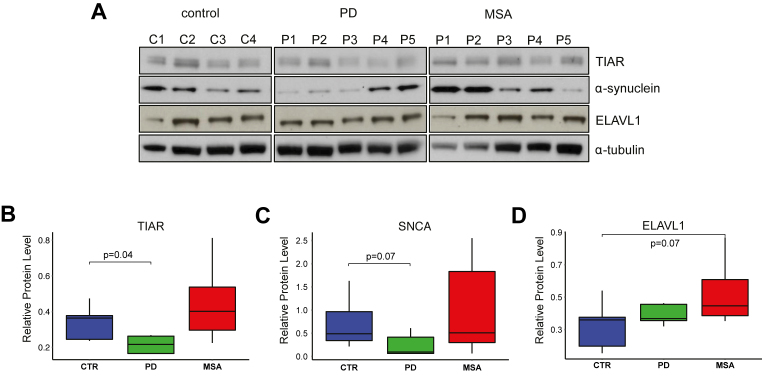
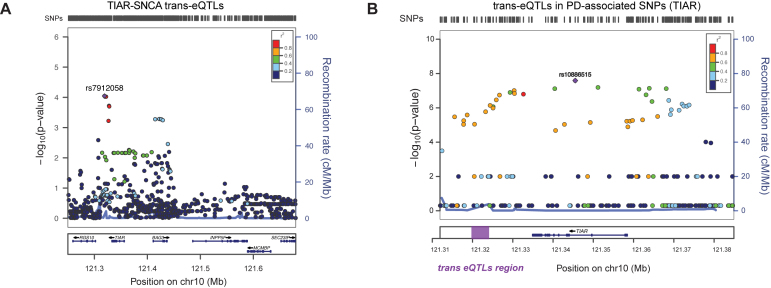
Recent evidence indicates a link between Parkinson's Disease (PD) and the expression of a-synuclein (SNCA) isoforms with different 3' untranslated regions (3'UTRs). Yet, the post-transcriptional mechanisms regulating SNCA expression are unknown. Using a large-scale in vitro /in silico screening we identified RNA-binding proteins (RBPs) that interact with SNCA 3' UTRs. We identified two RBPs, ELAVL1 and TIAR, that bind with high affinity to the most abundant and translationally active 3' UTR isoform (575 nt). Knockdown and overexpression experiments indicate that both ELAVL1 and TIAR positively regulate endogenous SNCA in vivo. The mechanism of regulation implies mRNA stabilization as well as enhancement of translation in the case of TIAR. We observed significant alteration of both TIAR and ELAVL1 expression in motor cortex of post-mortem brain donors and primary cultured fibroblast from patients affected by PD and Multiple System Atrophy (MSA). Moreover, trans expression quantitative trait loci (trans-eQTLs) analysis revealed that a group of single nucleotide polymorphisms (SNPs) in TIAR genomic locus influences SNCA expression in two different brain areas, nucleus accumbens and hippocampus. Our study sheds light on the 3' UTR-mediated regulation of SNCA and its link with PD pathogenesis, thus opening up new avenues for investigation of post-transcriptional mechanisms in neurodegeneration.
© The Author(s) 2017. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures
References
-
- de Lau L.M.L., Breteler M.M.B.. Epidemiology of Parkinson's disease. Lancet Neurol. 2006; 5:525–535. - PubMed
-
- Dauer W., Przedborski S.. Parkinson's disease: mechanisms and models. Neuron. 2003; 39:889–909. - PubMed
-
- Spillantini M.G., Schmidt M.L., Lee V.M., Trojanowski J.Q., Jakes R., Goedert M.. Alpha-synuclein in Lewy bodies. Nature. 1997; 388:839–840. - PubMed
-
- Zanzoni A., Marchese D., Agostini F., Bolognesi B., Cirillo D., Botta-Orfila M., Livi C.M., Rodriguez-Mulero S., Tartaglia G.G.. Principles of self-organization in biological pathways: a hypothesis on the autogenous association of alpha-synuclein. Nucleic Acids Res. 2013; 41:9987–9998. - PMC - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
