

Bias Factor and Therapeutic Window Correlate to Predict Safer Opioid Analgesics
- PMID: 29149605
- PMCID: PMC5731250
- DOI: 10.1016/j.cell.2017.10.035
Bias Factor and Therapeutic Window Correlate to Predict Safer Opioid Analgesics
Abstract
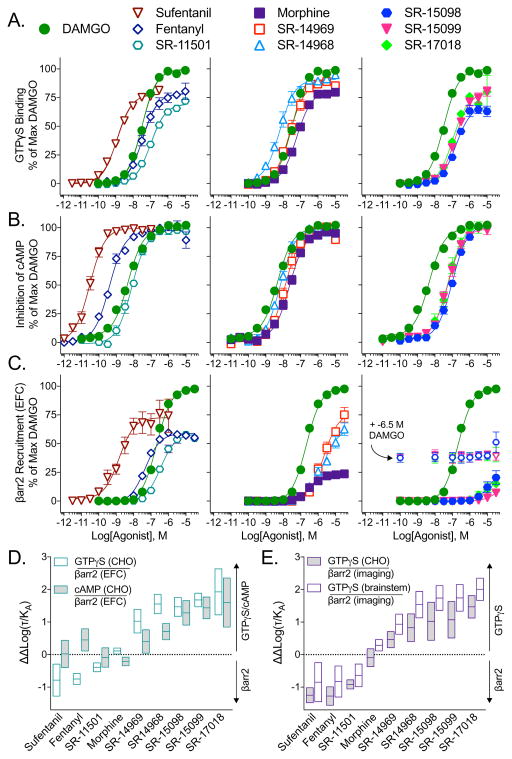
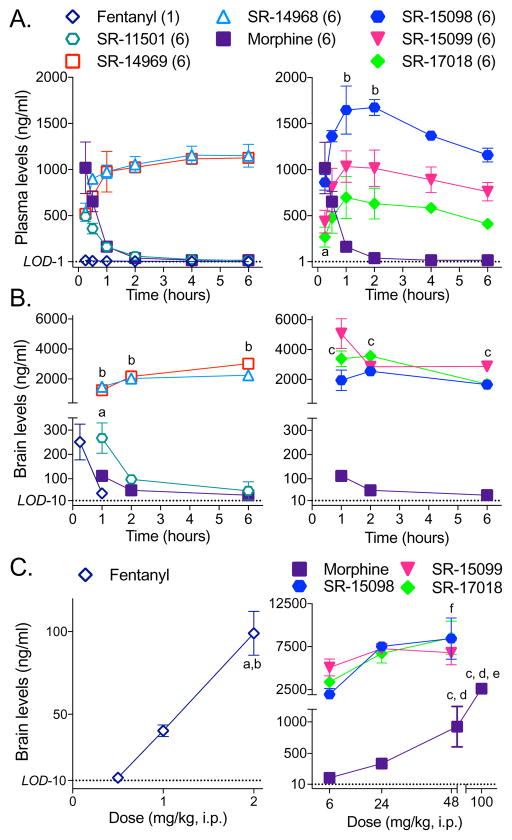
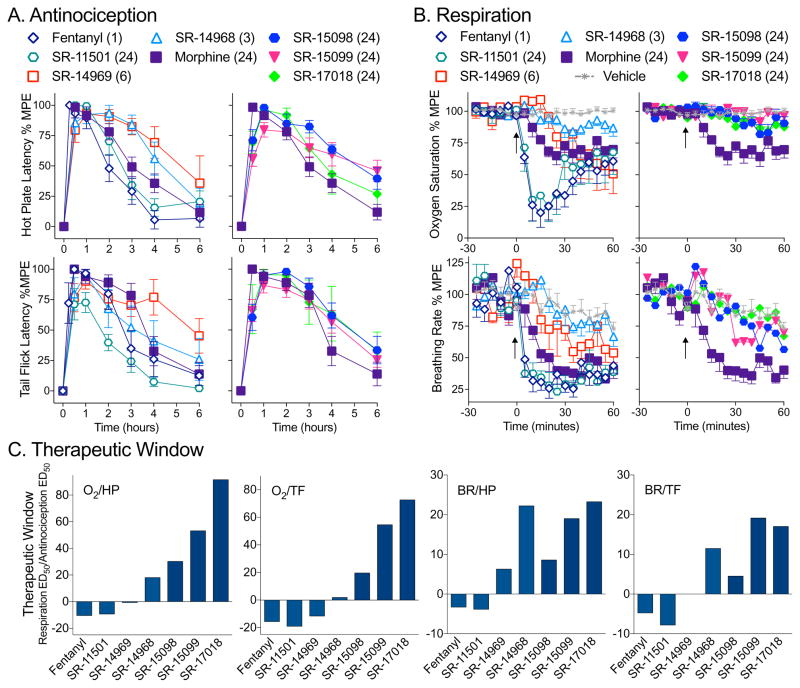
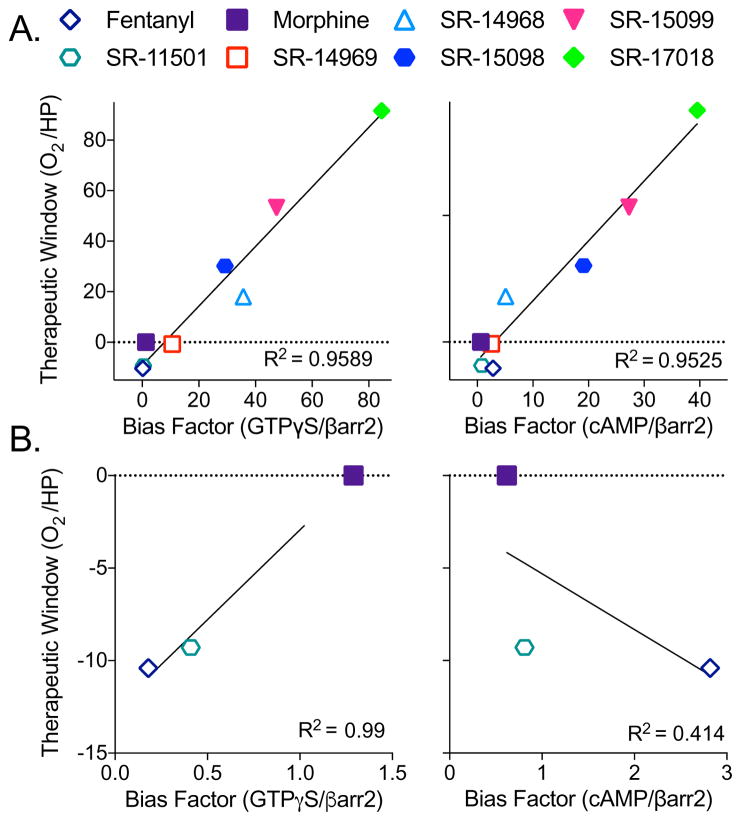
Biased agonism has been proposed as a means to separate desirable and adverse drug responses downstream of G protein-coupled receptor (GPCR) targets. Herein, we describe structural features of a series of mu-opioid-receptor (MOR)-selective agonists that preferentially activate receptors to couple to G proteins or to recruit βarrestin proteins. By comparing relative bias for MOR-mediated signaling in each pathway, we demonstrate a strong correlation between the respiratory suppression/antinociception therapeutic window in a series of compounds spanning a wide range of signaling bias. We find that βarrestin-biased compounds, such as fentanyl, are more likely to induce respiratory suppression at weak analgesic doses, while G protein signaling bias broadens the therapeutic window, allowing for antinociception in the absence of respiratory suppression.
Keywords: G protein-coupled receptor (GPCR); biased agonism; fentanyl; functional selectivity; morphine; mu opioid receptor; pain; respiration; side effects; βarrestin.
Copyright © 2017 Elsevier Inc. All rights reserved.
Figures
Comment in
-
Tuning Biased GPCR Signaling for Physiological Gain.Cell. 2017 Nov 16;171(5):989-991. doi: 10.1016/j.cell.2017.10.046. Cell. 2017. PMID: 29149613
-
Strategy for making safer opioids bolstered.Nature. 2018 Jan 18;553(7688):286-288. doi: 10.1038/d41586-018-00045-1. Nature. 2018. PMID: 29345640 No abstract available.
References
-
- Black JW, Leff P. Operational models of pharmacological agonism. Proc R Soc Lond B Biol Sci. 1983;220:141–162. - PubMed
-
- Bohn LM, Gainetdinov RR, Lin FT, Lefkowitz RJ, Caron MG. Mu-opioid receptor desensitization by beta-arrestin-2 determines morphine tolerance but not dependence. Nature. 2000;408:720–723. - PubMed
-
- Bohn LM, Lefkowitz RJ, Gainetdinov RR, Peppel K, Caron MG, Lin FT. Enhanced morphine analgesia in mice lacking beta-arrestin 2. Science. 1999;286:2495–2498. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
