

Chronic Stress Alters Striosome-Circuit Dynamics, Leading to Aberrant Decision-Making
- PMID: 29149606
- PMCID: PMC5734095
- DOI: 10.1016/j.cell.2017.10.017
Chronic Stress Alters Striosome-Circuit Dynamics, Leading to Aberrant Decision-Making
Abstract
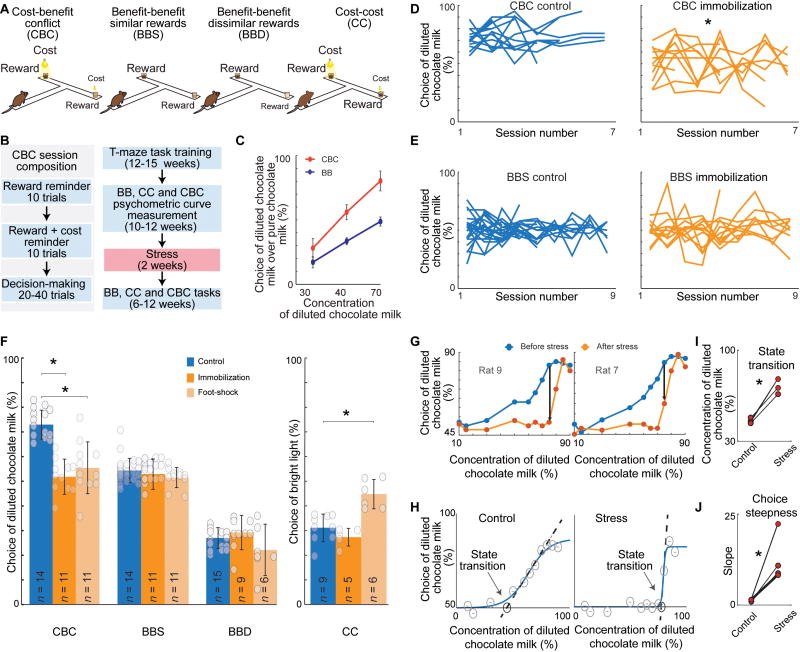
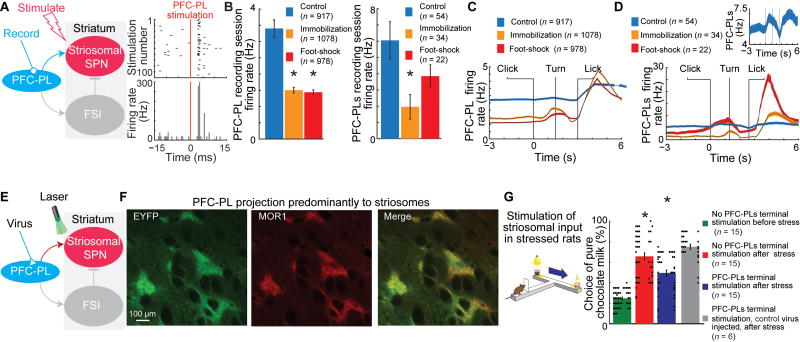
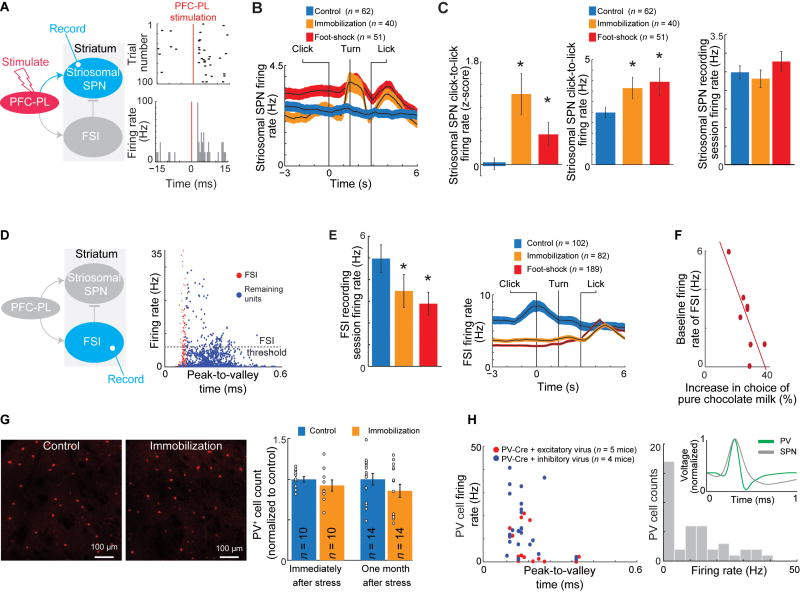
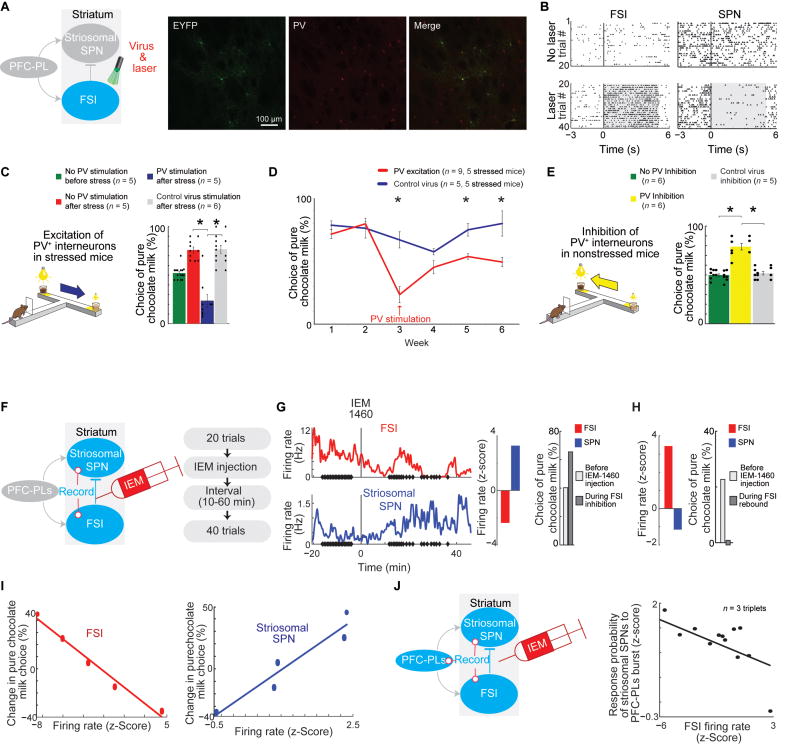
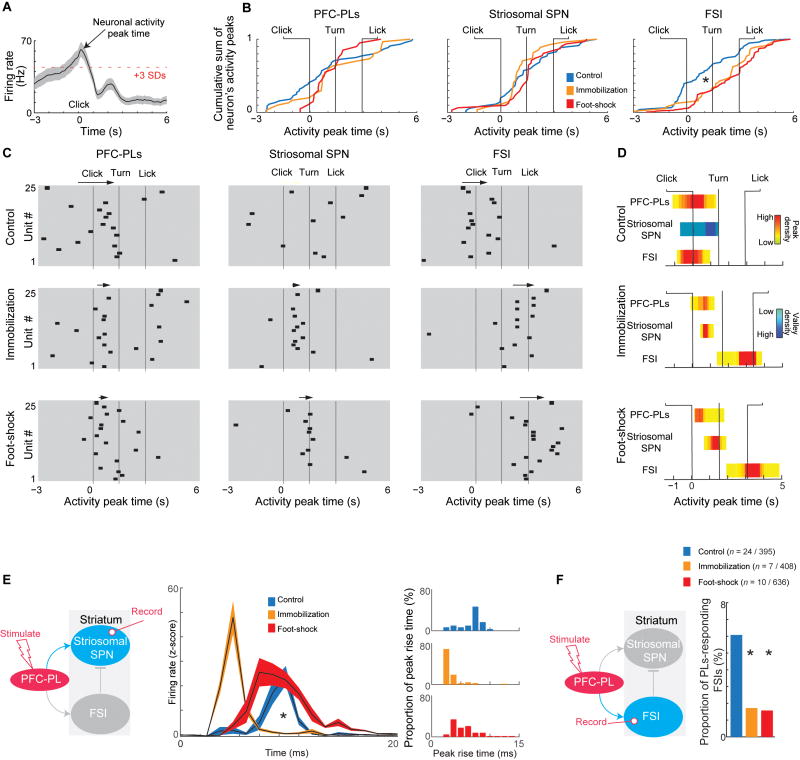
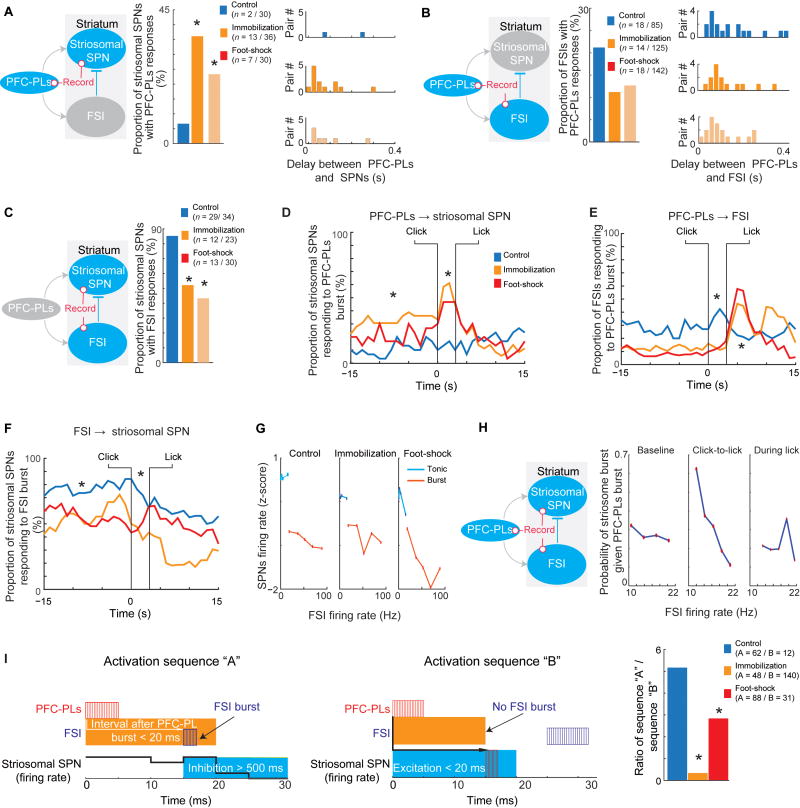
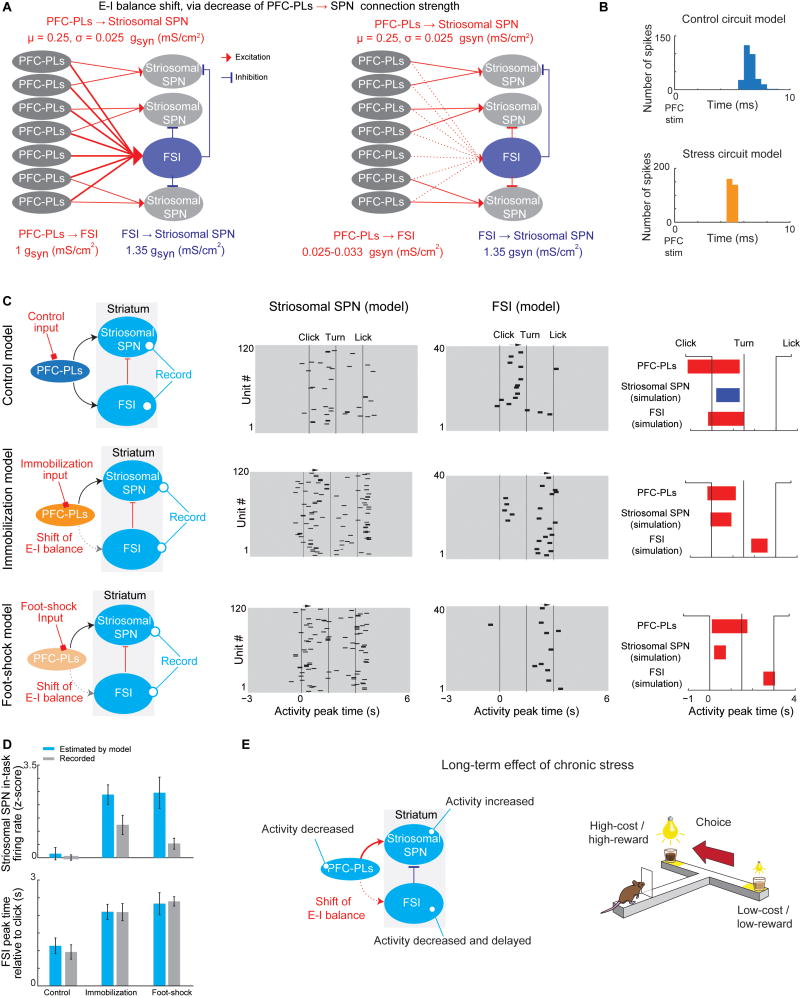
Effective evaluation of costs and benefits is a core survival capacity that in humans is considered as optimal, "rational" decision-making. This capacity is vulnerable in neuropsychiatric disorders and in the aftermath of chronic stress, in which aberrant choices and high-risk behaviors occur. We report that chronic stress exposure in rodents produces abnormal evaluation of costs and benefits resembling non-optimal decision-making in which choices of high-cost/high-reward options are sharply increased. Concomitantly, alterations in the task-related spike activity of medial prefrontal neurons correspond with increased activity of their striosome-predominant striatal projection neuron targets and with decreased and delayed striatal fast-firing interneuron activity. These effects of chronic stress on prefronto-striatal circuit dynamics could be blocked or be mimicked by selective optogenetic manipulation of these circuits. We suggest that altered excitation-inhibition dynamics of striosome-based circuit function could be an underlying mechanism by which chronic stress contributes to disorders characterized by aberrant decision-making under conflict. VIDEO ABSTRACT.
Keywords: basal ganglia; cost-benefit; excitation-inhibition balance; fast-spiking interneurons; optogenetics; parvalbumin-positive interneurons; prefrontal cortex; prelimbic cortex; striatum.
Copyright © 2017 Elsevier Inc. All rights reserved.
Figures
Comment in
-
Risky Business: The Circuits that Impact Stress-Induced Decision-Making.Cell. 2017 Nov 16;171(5):992-993. doi: 10.1016/j.cell.2017.11.004. Cell. 2017. PMID: 29149614
References
-
- Amat J, Baratta MV, Paul E, Bland ST, Watkins LR, Maier SF. Medial prefrontal cortex determines how stressor controllability affects behavior and dorsal raphe nucleus. Nat Neurosci. 2005;8:365–371. - PubMed
-
- Ampuero E, Luarte A, Santibanez M, Varas-Godoy M, Toledo J, Diaz-Veliz G, Cavada G, Rubio FJ, Wyneken U. Two chronic stress models based on movement restriction in rats respond selectively to antidepressant drugs: Aldolase c as a cotential biomarker. Int J Neuropsychopharmacol. 2015;8:pyv038. - PMC - PubMed
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
