

A cheZ-Like Gene in Azorhizobium caulinodans Is a Key Gene in the Control of Chemotaxis and Colonization of the Host Plant
- PMID: 29150498
- PMCID: PMC5772239
- DOI: 10.1128/AEM.01827-17
A cheZ-Like Gene in Azorhizobium caulinodans Is a Key Gene in the Control of Chemotaxis and Colonization of the Host Plant
Abstract
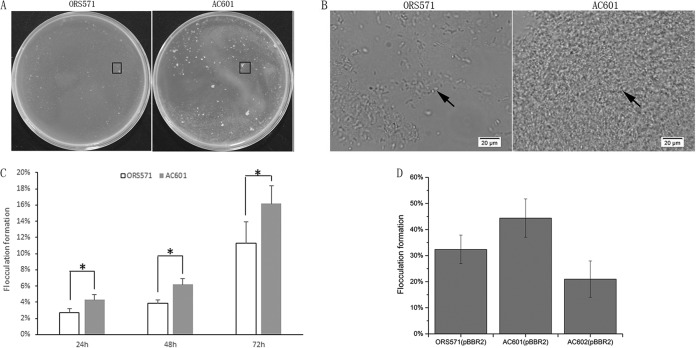
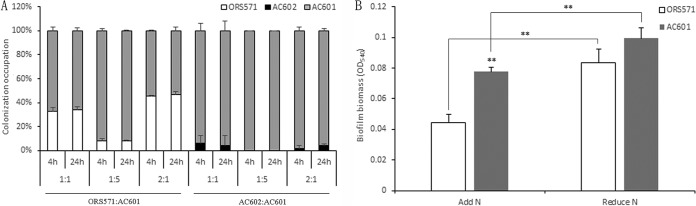
Chemotaxis can provide bacteria with competitive advantages for survival in complex environments. The CheZ chemotaxis protein is a phosphatase, affecting the flagellar motor in Escherichia coli by dephosphorylating the response regulator phosphorylated CheY protein (CheY∼P) responsible for clockwise rotation. A cheZ gene has been found in Azorhizobium caulinodans ORS571, in contrast to other rhizobial species studied so far. The CheZ protein in strain ORS571 has a conserved motif similar to that corresponding to the phosphatase active site in E. coli The construction of a cheZ deletion mutant strain and of cheZ mutant strains carrying a mutation in residues of the putative phosphatase active site showed that strain ORS571 participates in chemotaxis and motility, causing a hyperreversal behavior. In addition, the properties of the cheZ deletion mutant revealed that ORS571 CheZ is involved in other physiological processes, since it displayed increased flocculation, biofilm formation, exopolysaccharide (EPS) production, and host root colonization. In particular, it was observed that the expression of several exp genes, involved in EPS synthesis, was upregulated in the cheZ mutant compared to that in the wild type, suggesting that CheZ negatively controls exp gene expression through an unknown mechanism. It is proposed that CheZ influences the Azorhizobium-plant association by negatively regulating early colonization via the regulation of EPS production. This report established that CheZ in A. caulinodans plays roles in chemotaxis and the symbiotic association with the host plant.IMPORTANCE Chemotaxis allows bacteria to swim toward plant roots and is beneficial to the establishment of various plant-microbe associations. The level of CheY phosphorylation (CheY∼P) is central to the chemotaxis signal transduction. The mechanism of the signal termination of CheY∼P remains poorly characterized among Alphaproteobacteria, except for Sinorhizobium meliloti, which does not contain CheZ but which controls CheY∼P dephosphorylation through a phosphate sink mechanism. Azorhizobium caulinodans ORS571, a microsymbiont of Sesbania rostrata, has an orphan cheZ gene besides two cheY genes similar to those in S. meliloti In addition to controlling the chemotaxis response, the CheZ-like protein in strain ORS571 is playing a role by decreasing bacterial adhesion to the host plant, in contrast to the general situation where chemotaxis-associated proteins promote adhesion. In this study, we identified a CheZ-like protein among Alphaproteobacteria functioning in chemotaxis and the A. caulinodans-S. rostrata symbiosis.
Keywords: Azorhizobium caulinodans; CheZ; Sesbania rostrata; chemotaxis; colonization.
Copyright © 2018 American Society for Microbiology.
Figures





Similar articles
-
Inactivation of the Phosphatase CheZ Alters Cell-Surface Properties of Azorhizobium caulinodans ORS571 and Symbiotic Association with Sesbania rostrata.Mol Plant Microbe Interact. 2019 Nov;32(11):1547-1556. doi: 10.1094/MPMI-05-19-0143-R. Epub 2019 Sep 27. Mol Plant Microbe Interact. 2019. PMID: 31287368
-
CheY1 and CheY2 of Azorhizobium caulinodans ORS571 Regulate Chemotaxis and Competitive Colonization with the Host Plant.Appl Environ Microbiol. 2020 Jul 20;86(15):e00599-20. doi: 10.1128/AEM.00599-20. Print 2020 Jul 20. Appl Environ Microbiol. 2020. PMID: 32471918 Free PMC article.
-
Azorhizobium caulinodans Chemotaxis Is Controlled by an Unusual Phosphorelay Network.J Bacteriol. 2022 Feb 15;204(2):e0052721. doi: 10.1128/JB.00527-21. Epub 2021 Nov 29. J Bacteriol. 2022. PMID: 34843377 Free PMC article.
-
Control of bacterial chemotaxis.Mol Microbiol. 1996 Jun;20(5):903-10. doi: 10.1111/j.1365-2958.1996.tb02531.x. Mol Microbiol. 1996. PMID: 8809743 Review.
-
Bacterial tactic responses.Adv Microb Physiol. 1999;41:229-89. doi: 10.1016/s0065-2911(08)60168-x. Adv Microb Physiol. 1999. PMID: 10500847 Review.
Cited by
-
The effect of Azorhizobium caulinodans ORS571 and γ-aminobutyric acid on salt tolerance of Sesbania rostrata.Front Plant Sci. 2022 Aug 5;13:926850. doi: 10.3389/fpls.2022.926850. eCollection 2022. Front Plant Sci. 2022. PMID: 36046585 Free PMC article.
-
Rhizobial Chemotaxis and Motility Systems at Work in the Soil.Front Plant Sci. 2021 Aug 27;12:725338. doi: 10.3389/fpls.2021.725338. eCollection 2021. Front Plant Sci. 2021. PMID: 34512702 Free PMC article. Review.
-
Endophytism: A Multidimensional Approach to Plant-Prokaryotic Microbe Interaction.Front Microbiol. 2022 May 12;13:861235. doi: 10.3389/fmicb.2022.861235. eCollection 2022. Front Microbiol. 2022. PMID: 35633681 Free PMC article. Review.
-
Effects of Calcium and Signal Sensing Systems on Azorhizobium caulinodans Biofilm Formation and Host Colonization.Front Microbiol. 2020 Sep 16;11:563367. doi: 10.3389/fmicb.2020.563367. eCollection 2020. Front Microbiol. 2020. PMID: 33072026 Free PMC article.
-
Systematic Analysis of Lysine Acetylation Reveals Diverse Functions in Azorhizobium caulinodans Strain ORS571.Microbiol Spectr. 2023 Feb 14;11(1):e0353922. doi: 10.1128/spectrum.03539-22. Epub 2022 Dec 8. Microbiol Spectr. 2023. PMID: 36475778 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
