

Divalent Metal Cations Potentiate the Predatory Capacity of Amoeba for Cryptococcus neoformans
- PMID: 29150507
- PMCID: PMC5772251
- DOI: 10.1128/AEM.01717-17
Divalent Metal Cations Potentiate the Predatory Capacity of Amoeba for Cryptococcus neoformans
Abstract
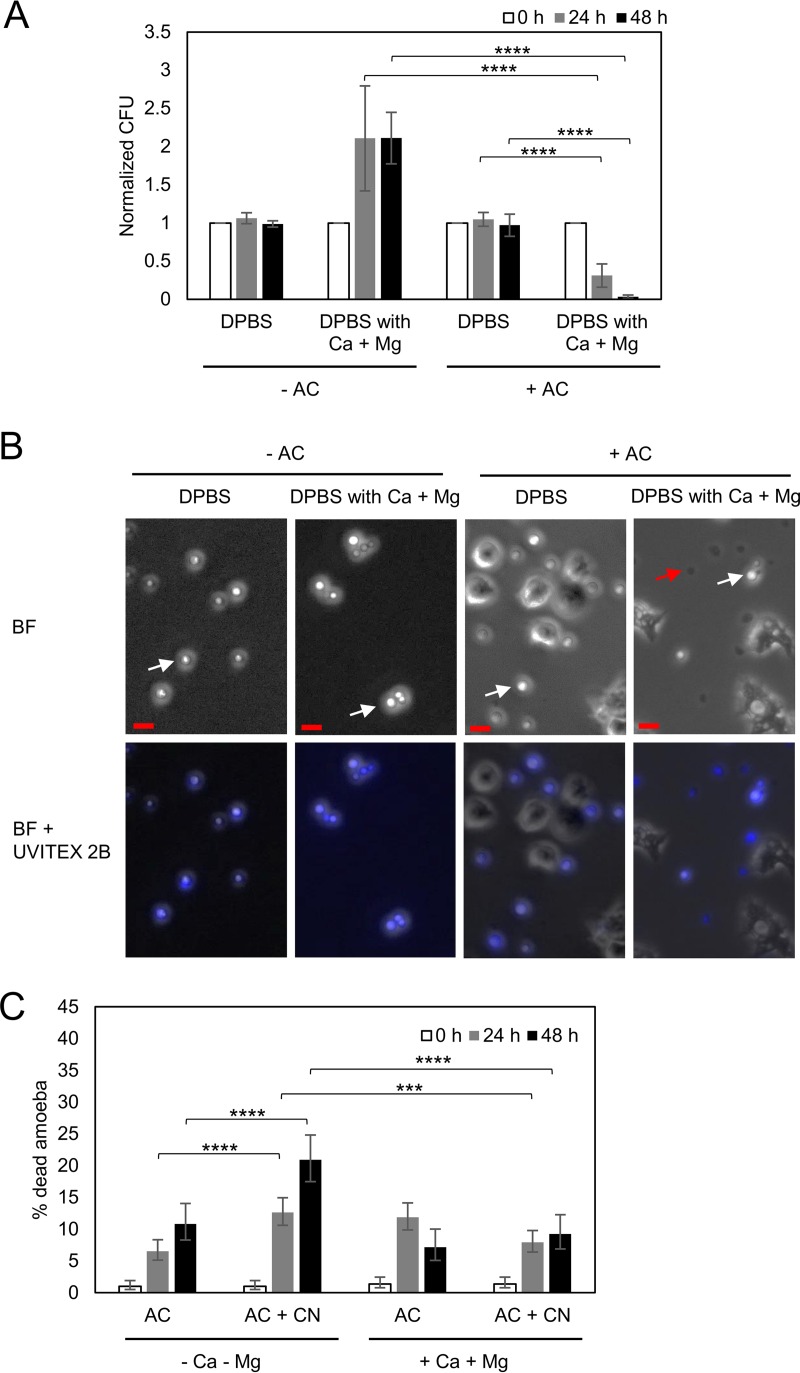
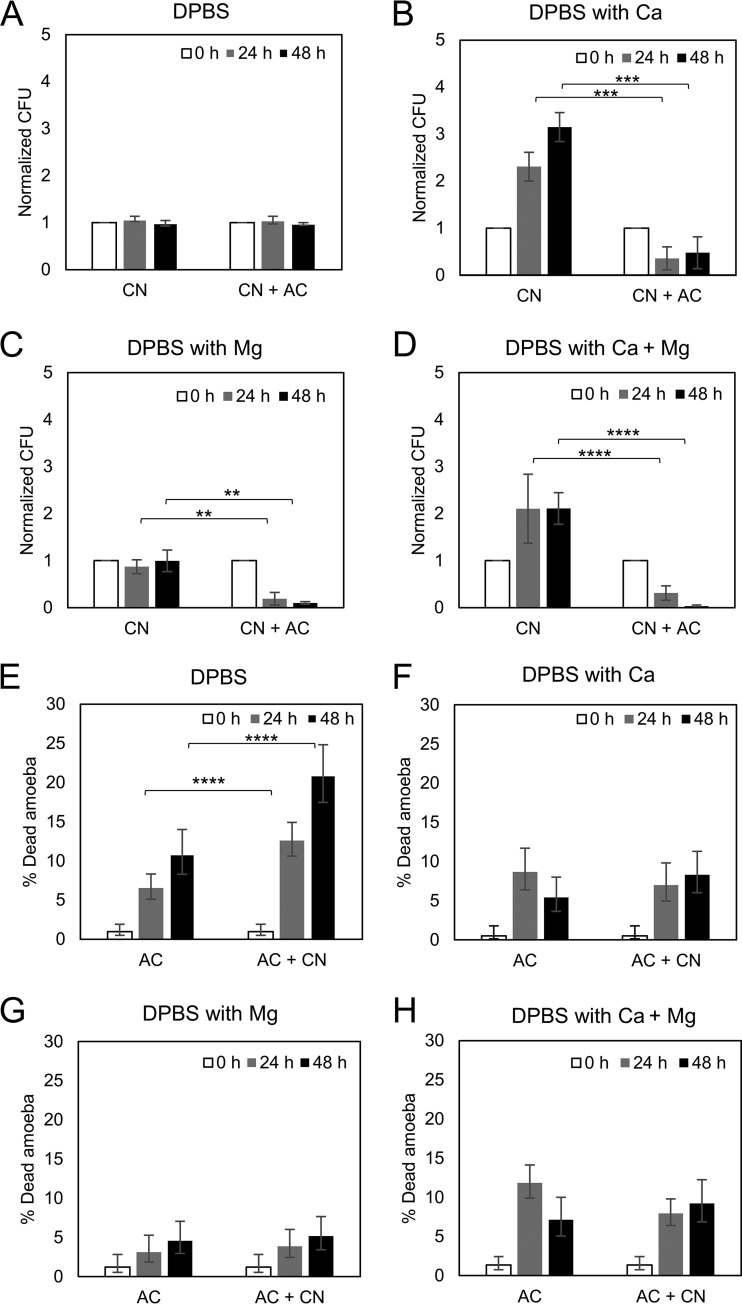
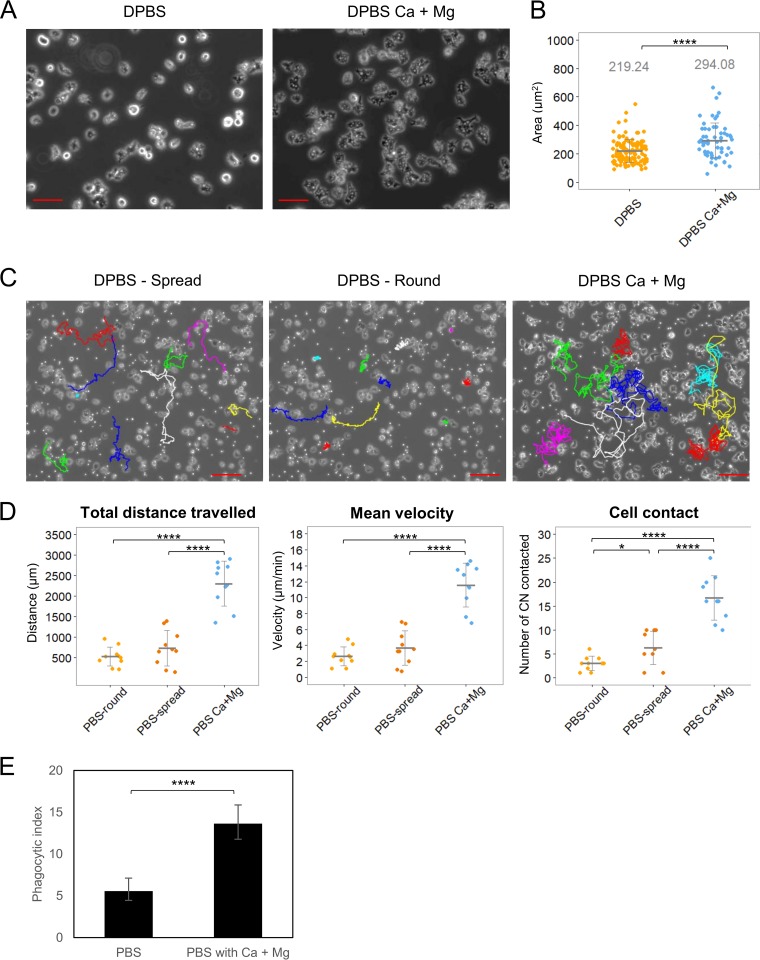
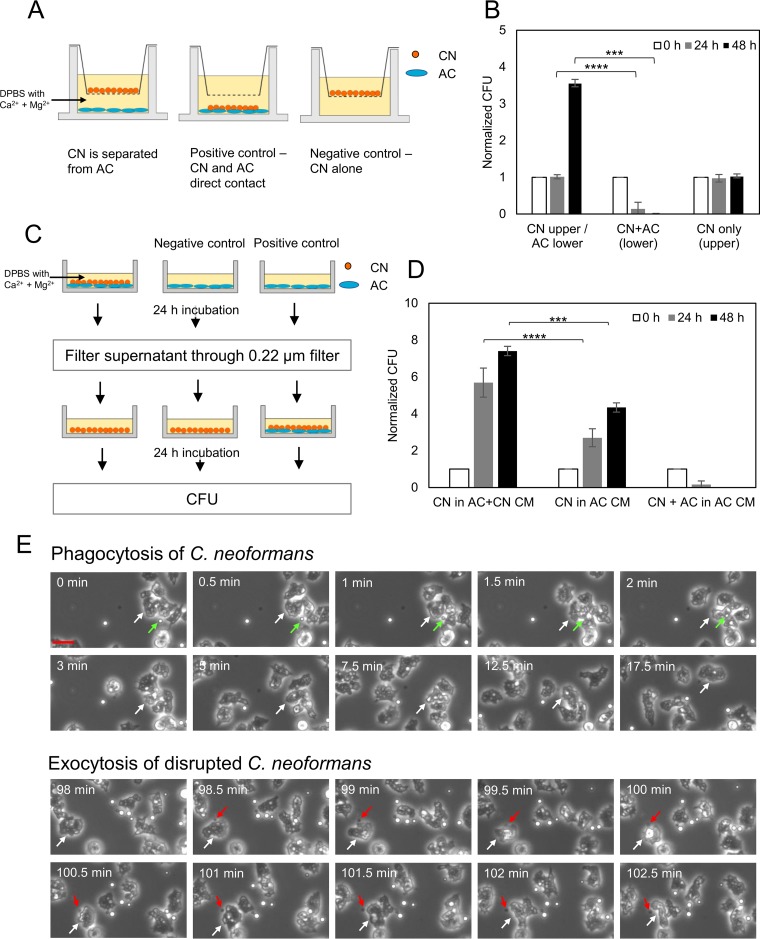
Among the best-studied interactions between soil phagocytic predators and a human-pathogenic fungus is that of Acanthamoeba castellanii and Cryptococcus neoformans The experimental conditions used in amoeba-fungus confrontation assays can have major effects on whether the fungus or the protozoan is ascendant in the interaction. In the presence of Mg2+ and Ca2+ in phosphate-buffered saline (PBS), C. neoformans was consistently killed when incubated with A. castellaniiA. castellanii survived better in the presence of Mg2+ and Ca2+, even when incubated with C. neoformans In the absence of Mg2+ and Ca2+, C. neoformans survived when incubated with A. castellanii, and the percentage of dead amoebae was higher than when incubated without yeast cells. These results show that the presence of Mg2+ and Ca2+ can make a decisive contribution toward tilting the outcome of the interaction in favor of the amoeba. Of the two metals, Mg2+ had a stronger effect than Ca2+ The cations enhanced A. castellanii activity against C. neoformans via enhanced phagocytosis, which is the major mechanism by which amoebae kill fungal cells. We found no evidence that amoebae use extracellular killing mechanisms in their interactions with C. neoformans In summary, the presence of Mg2+ and Ca2+ enhanced the cell adhesion on the surfaces and the motility of the amoeba, thus increasing the chance for contact with C. neoformans and the frequency of phagocytosis. Our findings imply that the divalent cation concentration in soils could be an important variable for whether amoebae can control C. neoformans in the environment.IMPORTANCE The grazing of soil organisms by phagocytic predators such as amoebae is thought to select for traits that enable some of them to acquire the capacity for virulence in animals. Consequently, knowledge about the interactions between amoebae and soil microbes, such as pathogenic fungi, is important for understanding how virulence can emerge. We show that the interaction between an amoeba and the pathogenic fungus C. neoformans is influenced by the presence in the assay of magnesium and calcium, which potentiate amoebae. The results may also have practical applications, since enriching soils with divalent cations may reduce C. neoformans numbers in contaminated soils.
Keywords: Cryptococcus neoformans; amoeba; cations; fungi; predation.
Copyright © 2018 American Society for Microbiology.
Figures
Similar articles
-
Amoeba Predation of Cryptococcus neoformans Results in Pleiotropic Changes to Traits Associated with Virulence.mBio. 2021 Apr 27;12(2):e00567-21. doi: 10.1128/mBio.00567-21. mBio. 2021. PMID: 33906924 Free PMC article.
-
Phagocytosis of Cryptococcus neoformans by, and nonlytic exocytosis from, Acanthamoeba castellanii.Appl Environ Microbiol. 2010 Sep;76(18):6056-62. doi: 10.1128/AEM.00812-10. Epub 2010 Jul 30. Appl Environ Microbiol. 2010. PMID: 20675457 Free PMC article.
-
Cryptococcus neoformans interactions with amoebae suggest an explanation for its virulence and intracellular pathogenic strategy in macrophages.Proc Natl Acad Sci U S A. 2001 Dec 18;98(26):15245-50. doi: 10.1073/pnas.261418798. Epub 2001 Dec 11. Proc Natl Acad Sci U S A. 2001. PMID: 11742090 Free PMC article.
-
Host-pathogen interactions: lessons from phagocytic predation on fungi.Curr Opin Microbiol. 2021 Aug;62:38-44. doi: 10.1016/j.mib.2021.04.010. Epub 2021 May 26. Curr Opin Microbiol. 2021. PMID: 34051610 Review.
-
Amoeba provide insight into the origin of virulence in pathogenic fungi.Adv Exp Med Biol. 2012;710:1-10. doi: 10.1007/978-1-4419-5638-5_1. Adv Exp Med Biol. 2012. PMID: 22127880 Review.
Cited by
-
A hidden battle in the dirt: Soil amoebae interactions with Paracoccidioides spp.PLoS Negl Trop Dis. 2019 Oct 7;13(10):e0007742. doi: 10.1371/journal.pntd.0007742. eCollection 2019 Oct. PLoS Negl Trop Dis. 2019. PMID: 31589617 Free PMC article.
-
The 'Amoeboid Predator-Fungal Animal Virulence' Hypothesis.J Fungi (Basel). 2019 Jan 21;5(1):10. doi: 10.3390/jof5010010. J Fungi (Basel). 2019. PMID: 30669554 Free PMC article. Review.
-
Similar evolutionary trajectories in an environmental Cryptococcus neoformans isolate after human and murine infection.Proc Natl Acad Sci U S A. 2023 Jan 10;120(2):e2217111120. doi: 10.1073/pnas.2217111120. Epub 2023 Jan 5. Proc Natl Acad Sci U S A. 2023. PMID: 36603033 Free PMC article.
-
Human Conjunctival Transcriptome in Acanthamoeba Keratitis: An Exploratory Study.Cornea. 2024 Oct 1;43(10):1272-1277. doi: 10.1097/ICO.0000000000003545. Epub 2024 May 21. Cornea. 2024. PMID: 38771726
-
The enigmatic role of fungal annexins: the case of Cryptococcus neoformans.Microbiology (Reading). 2019 Aug;165(8):852-862. doi: 10.1099/mic.0.000815. Epub 2019 May 29. Microbiology (Reading). 2019. PMID: 31140968 Free PMC article.
References
-
- Casadevall A, Perfect JR. 1998. Cryptococcus neoformans. American Society for Microbiology, Washington, DC.
-
- Casadevall A, Nosanchuk JD, Steenbergen JN. 2003. “Ready-made” virulence and “dual-use” virulence factors in pathogenic environmental fungi–the Cryptococcus neoformans paradigm. Curr Opin Microbiol 112:1164–1175. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
