

Cancer cells induce interleukin-22 production from memory CD4+ T cells via interleukin-1 to promote tumor growth
- PMID: 29150554
- PMCID: PMC5724250
- DOI: 10.1073/pnas.1705165114
Cancer cells induce interleukin-22 production from memory CD4+ T cells via interleukin-1 to promote tumor growth
Abstract
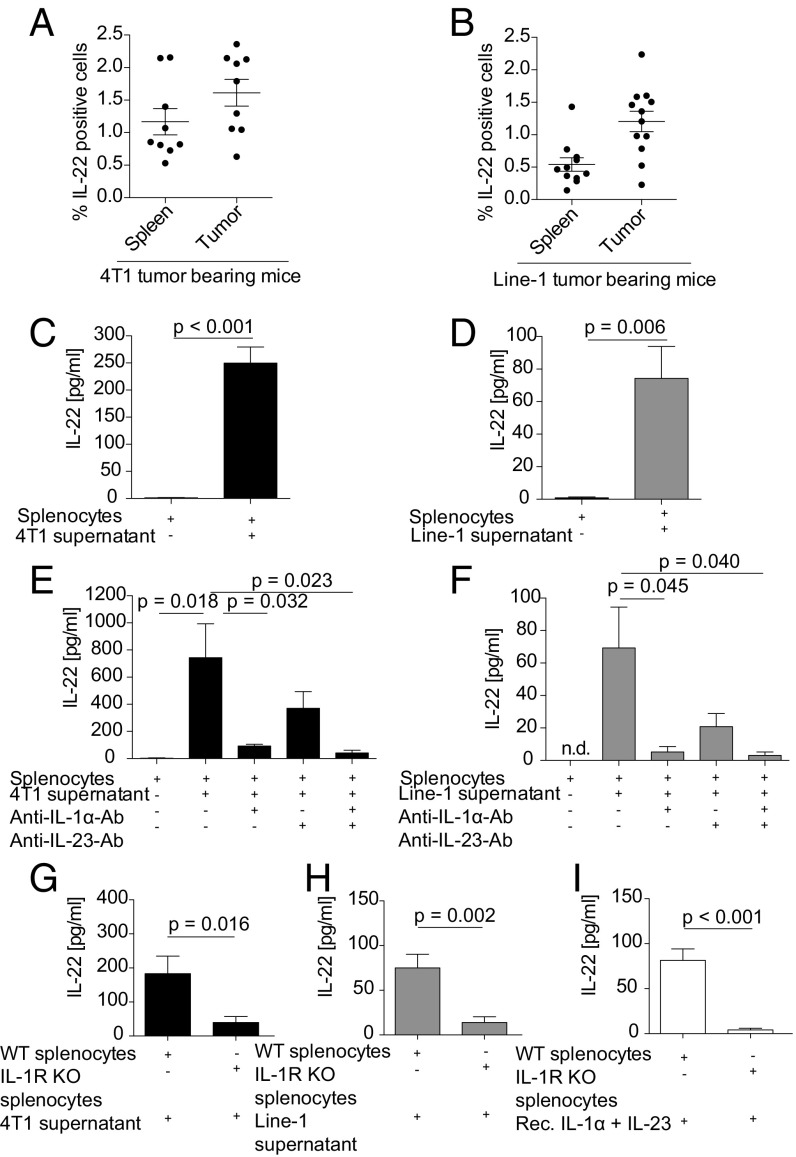
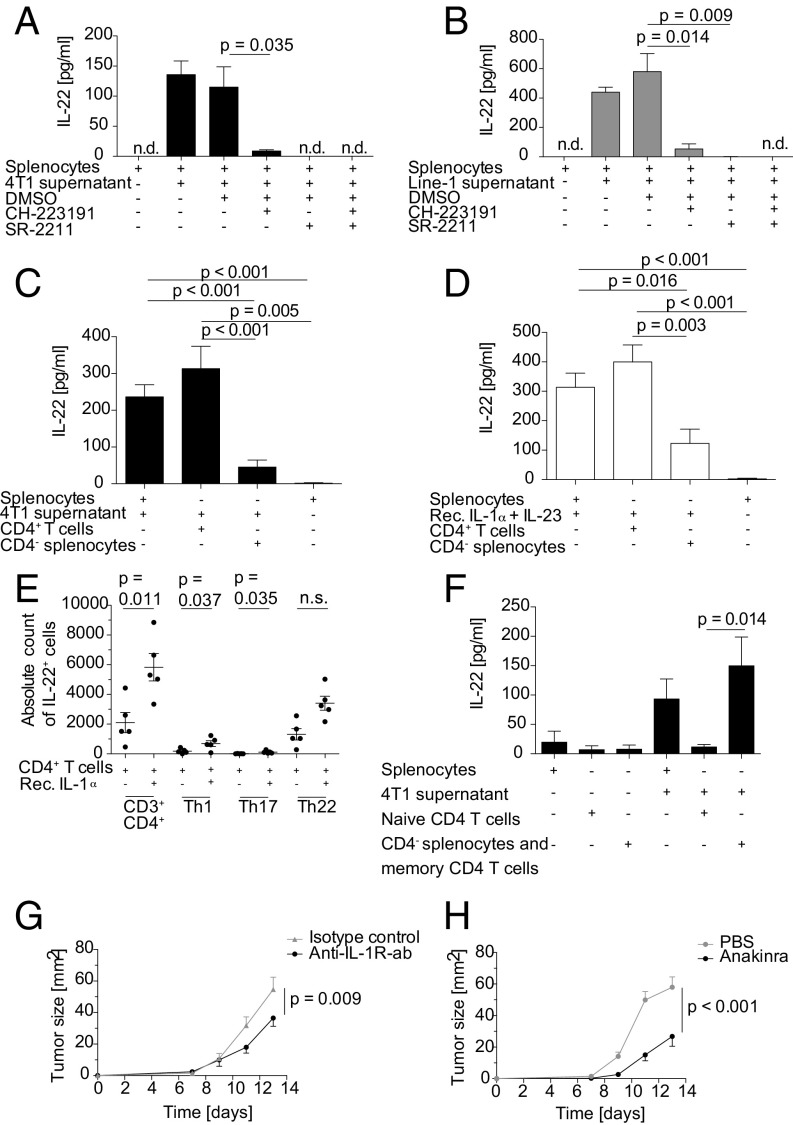
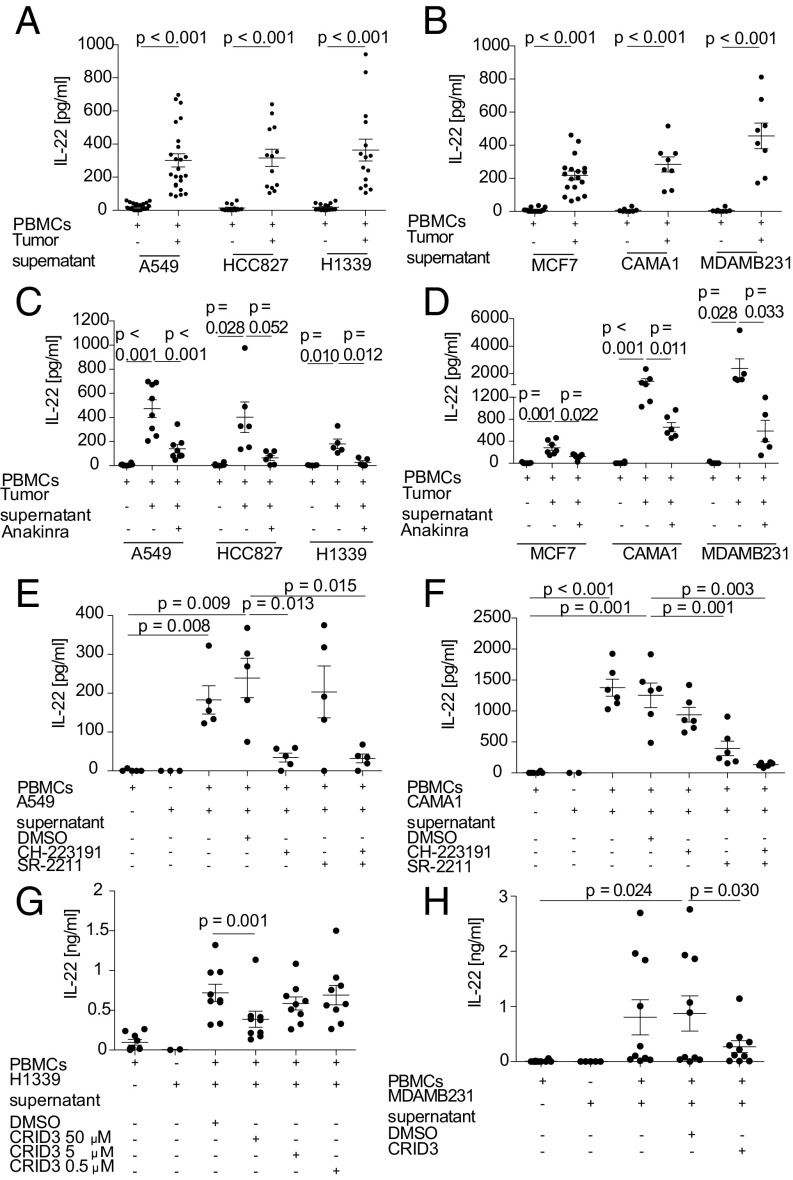
IL-22 has been identified as a cancer-promoting cytokine that is secreted by infiltrating immune cells in several cancer models. We hypothesized that IL-22 regulation would occur at the interface between cancer cells and immune cells. Breast and lung cancer cells of murine and human origin induced IL-22 production from memory CD4+ T cells. In the present study, we found that IL-22 production in humans is dependent on activation of the NLRP3 inflammasome with the subsequent release of IL-1β from both myeloid and T cells. IL-1 receptor signaling via the transcription factors AhR and RORγt in T cells was necessary and sufficient for IL-22 production. In these settings, IL-1 induced IL-22 production from a mixed T helper cell population comprised of Th1, Th17, and Th22 cells, which was abrogated by the addition of anakinra. We confirmed these findings in vitro and in vivo in two murine tumor models, in primary human breast and lung cancer cells, and in deposited expression data. Relevant to ongoing clinical trials in breast cancer, we demonstrate here that the IL-1 receptor antagonist anakinra abrogates IL-22 production and reduces tumor growth in a murine breast cancer model. Thus, we describe here a previously unrecognized mechanism by which cancer cells induce IL-22 production from memory CD4+ T cells via activation of the NLRP3 inflammasome and the release of IL-1β to promote tumor growth. These findings may provide the basis for therapeutic interventions that affect IL-22 production by targeting IL-1 activity.
Keywords: anakinra; cancer immunology; inflammasome; interleukin-1; interleukin-22.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Lim C, Savan R. The role of the IL-22/IL-22R1 axis in cancer. Cytokine Growth Factor Rev. 2014;25:257–271. - PubMed
-
- Hanahan D, Weinberg RA. Hallmarks of cancer: The next generation. Cell. 2011;144:646–674. - PubMed
-
- Sonnenberg GF, Fouser LA, Artis D. Border patrol: Regulation of immunity, inflammation and tissue homeostasis at barrier surfaces by IL-22. Nat Immunol. 2011;12:383–390.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
