

Review
doi: 10.1104/pp.17.01415.
Epub 2017 Nov 17.
ROP GTPases Structure-Function and Signaling Pathways
Affiliations
- PMID: 29150557
- PMCID: PMC5761820
- DOI: 10.1104/pp.17.01415
Item in Clipboard
Review
ROP GTPases Structure-Function and Signaling Pathways
Plant Physiol.
2018 Jan.
Abstract
Interactions between receptor like kinases and guanyl nucleotide exchange factors together with identification of effector proteins reveal putative ROP GTPases signaling cascades.
Figures
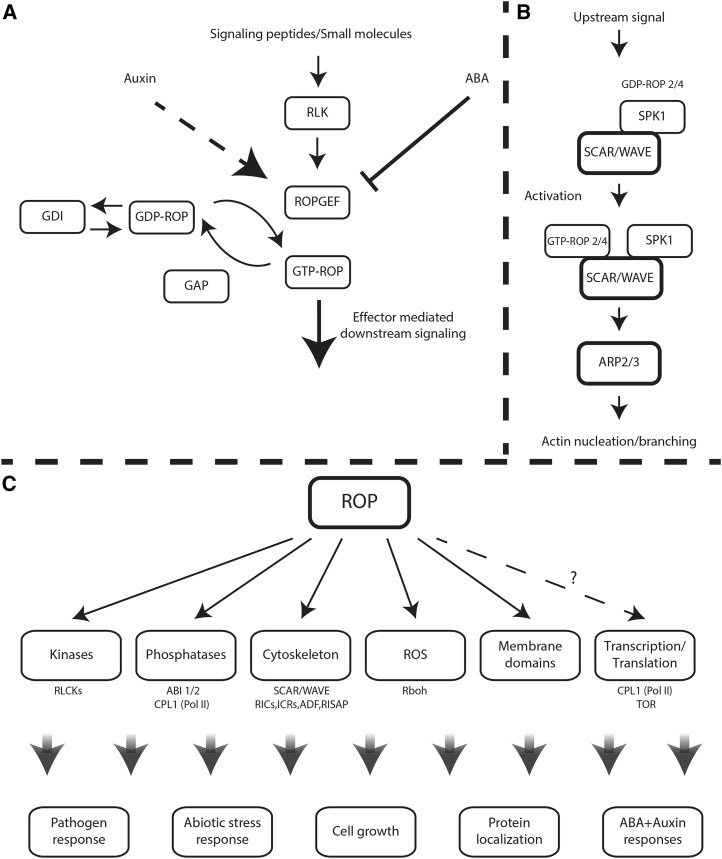
ROP signaling pathways. A, ROP activation/deactivation cycles. ROPs cycle between a GTP-bound active state and a GDP-bound inactive state. Activation is regulated by ROP-specific GEFs, inactivation is enhanced by ROP-specific GAPs, and recycling of type I ROPs between the cytosol and the plasma membrane is facilitated by RhoGDIs. Some GEFs are activated by peptide-activated, plasma membrane-associated RLKs. ROPs also are up-regulated by auxin by an as yet unknown pathway (dashed arrow) and inactivated by ABA, which enhances the degradation of ROPGEFs. B, The Dock family GEF homolog SPK1 is associated with the WAVE complex. Upon activation, ROP-GTP interacts with the WAVE SRA1 subunit and possibly other subunits, resulting in WAVE activation. Activated WAVE activates actin nucleation/branching induced by Arp2/3. C, ROP downstream signaling. ROP regulated pathways and known effectors. The dashed arrow indicates uncertainty regarding the cellular targets of ROP. CPL1 is regulated by ROP signaling but does not interact physically with ROPs. TOR interacts weakly with GTP-bound ROP2 and more strongly with the GDP-bound form.
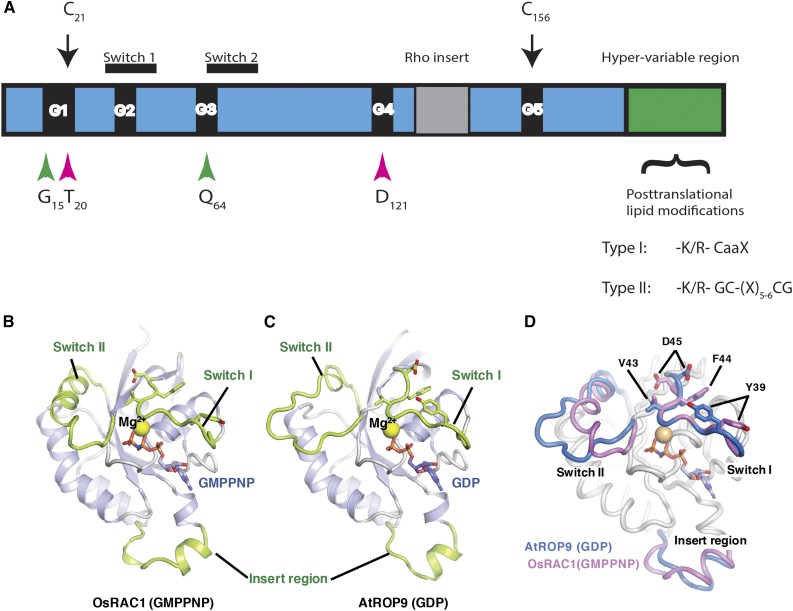
Structures of ROPs. A, Schematic model highlighting the conserved G-box motifs (G1–G5), switch I and II domains, the insert region, and the HVR. The positions of the conserved residues mutated in the constitutively active and dominant-negative mutations are highlighted by green and magenta arrowheads, respectively. C21 and C156 are conserved G domain Cys residues that undergo activation-dependent transient S-acylation (black arrows). The HVR terminates with a canonical CaaX box geranylgeranylation motif and a PBR in type I ROPs or with a GC-CG box and a proximal PBR in type II ROPs. B, Three-dimensional structure of GTP-bound OsRAC1 [OsRAC1 (GMPPNP)]. C, Three-dimensional structure of the GDP-bound AtROP9 [AtROP9 (GDP)]. In B and C, the switch I, switch II, and insert regions are colored green. GMPPNP and GDP are shown as stick models (red, oxygen; blue, nitrogen; orange, phosphorus). The Mg2+ ions are shown as yellow spheres. D, Superimposed structures of OsRAC1 (GMPPNP) and AtROP9 (GDP). The main chains of OsRAC1 (GMPPNP) and AtROP9 (GDP) (chain B; Protein Data Bank code 2J0V) were superimposed using PyMOL. Regions of the OsRAC1 and AtROP9 proteins are colored blue and pink, respectively. The side chains of four key switch I residues in OsRAC1 (Val-43, Phe-44, Asp-45, and Tyr-39) and equivalent residues in AtROP9 (Val-39, Phe-40, Asp-41, and Tyr-35) are shown in stick representation. B to D were prepared and contributed by Izuru Ohki and Chojiro Kojima and are based on their article describing the structure of GTP-bound OsRAC1 (Kosami et al., 2014).
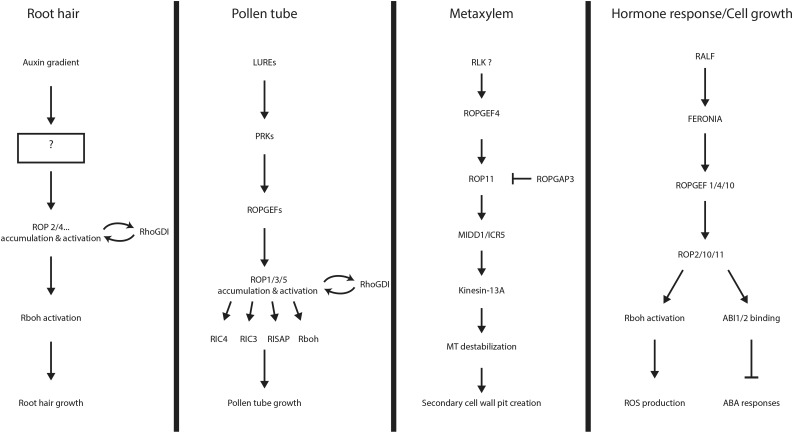
ROP signaling pathways. A schematic overview of identified ROP signaling pathways is shown. Components of the signaling pathways mentioned in the text are noted, and their relationships to each other are indicated by arrows. The question mark notes an area of insufficient data. From top to bottom in each section are listed the upstream signal, the receptors that relay the signal into the cell, the regulation of ROP activity, and the downstream effectors that modulate the cellular response.
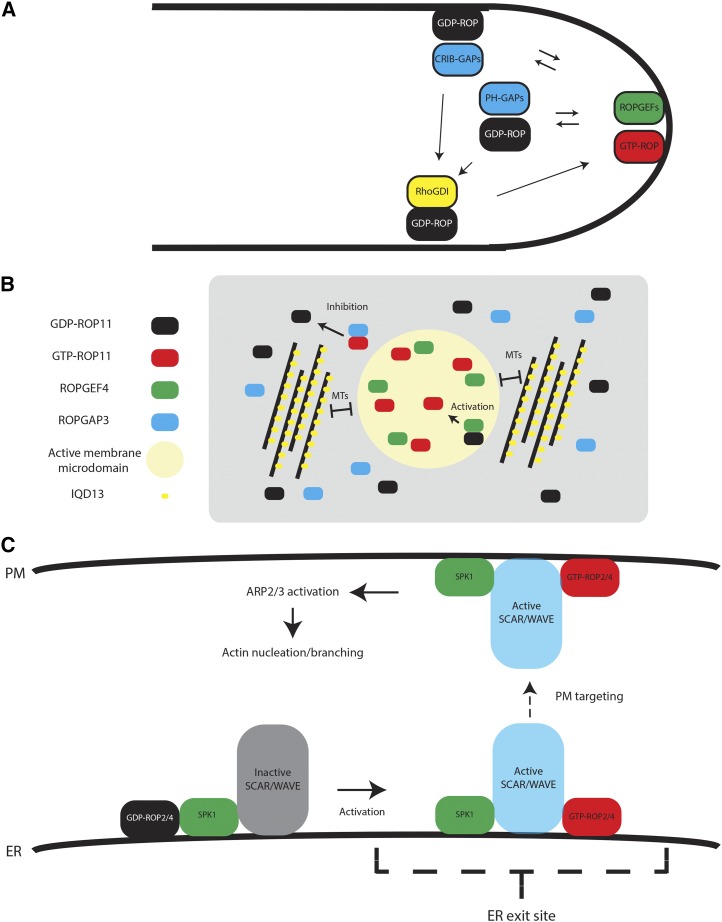
Mechanisms of spatial ROP activity regulation. A, Local activation of ROPs in the pollen tube. ROPs are activated at the tip by the activation of ROPGEFs by associated RLKs. CRIB domain ROPGAPs inactivate ROPs at the shank to maintain polar growth. PH domain GAPs are localized in subapical cytoplasmic vesicles, restricting ROP active domains. RhoGDIs recycle ROPs from the membrane. B, Formation of active ROP domains in differentiating metaxylem. During metaxylem differentiation, ROP11 is coexpressed with ROPGEF4 and ROPGAP3, resulting in the formation of active ROP11 domains and global inactivation. The size of the ROP11 domains is restricted by MTs and the MT- and plasma membrane-associated protein IQD13. C, Activation of ROPs by SPK1 and the formation of active actin nucleation/branching domains. SPK1 is associated with the WAVE complex. Activation of ROPs by SPK1 causes their association with the SRA1 subunit of the WAVE complex and its activation. Active WAVE can then activate actin nucleation/branching by Arp2/3. ER, Endoplasmic reticulum; PM, plasma membrane.

ROP regulation of cytoskeleton organization. A, Cortical MT reordering by ROP6, RIC1, and KTN1. RIC1 interacts physically with and promotes the MT severing by KTN1, leading to MT reordering. B, Localized cortical MT depolymerization. In developing metaxylem, MT depolarization takes place by the recruitment of MIDD1 and Kinesin13A to active ROP11 domains. C, Actin dynamics in tip-growing pollen tubes. ROP regulates actin assembly by two counteracting pathways. RIC4 promotes assembly, whereas RIC3 causes disassembly by stimulating Ca2+ influx into the cytoplasm. D, Actin nucleation/branching in trichomes and pavement cells. ROPs and the ROPGEF SPK1 interact with the SCAR/WAVE complex and promote actin filament nucleation/branching.




Similar articles
-
The cellular function of ROP GTPase prenylation is important for multicellularity in the moss Physcomitrium patens.Development. 2022 Jun 15;149(12):dev200279. doi: 10.1242/dev.200279. Epub 2022 Jun 23. Development. 2022. PMID: 35660859
-
Small GTPases in vesicle trafficking.Curr Opin Plant Biol. 2004 Dec;7(6):694-700. doi: 10.1016/j.pbi.2004.09.014. Curr Opin Plant Biol. 2004. PMID: 15491918 Review.
-
ROP/RAC GTPase: an old new master regulator for plant signaling.Curr Opin Plant Biol. 2004 Oct;7(5):527-36. doi: 10.1016/j.pbi.2004.07.006. Curr Opin Plant Biol. 2004. PMID: 15337095 Review.
-
GTPases: a family of molecular switches and clocks.Philos Trans R Soc Lond B Biol Sci. 1995 Sep 29;349(1329):283-9. doi: 10.1098/rstb.1995.0114. Philos Trans R Soc Lond B Biol Sci. 1995. PMID: 8577839 Review.
-
ROP/RAC GTPase signaling.Curr Opin Plant Biol. 2007 Oct;10(5):490-4. doi: 10.1016/j.pbi.2007.07.005. Epub 2007 Aug 20. Curr Opin Plant Biol. 2007. PMID: 17709276 Free PMC article. Review.
Cited by
-
Microtubule-associated phase separation of MIDD1 tunes cell wall spacing in xylem vessels in Arabidopsis thaliana.Nat Plants. 2024 Jan;10(1):100-117. doi: 10.1038/s41477-023-01593-9. Epub 2024 Jan 3. Nat Plants. 2024. PMID: 38172572
-
Capsicum Leaves under Stress: Using Multi-Omics Analysis to Detect Abiotic Stress Network of Secondary Metabolism in Two Species.Antioxidants (Basel). 2022 Mar 30;11(4):671. doi: 10.3390/antiox11040671. Antioxidants (Basel). 2022. PMID: 35453356 Free PMC article.
-
ZmRop1 participates in maize defense response to the damage of Spodoptera frugiperda larvae through mediating ROS and soluble phenol production.Plant Direct. 2022 Dec 15;6(12):e468. doi: 10.1002/pld3.468. eCollection 2022 Dec. Plant Direct. 2022. PMID: 36540415 Free PMC article.
-
Abscisic acid switches cell division modes of asymmetric cell division and symmetric cell division in stem cells of protonemal filaments in the moss Physcomitrium patens.Plant Biotechnol (Tokyo). 2022 Mar 25;39(1):13-17. doi: 10.5511/plantbiotechnology.22.0107a. Plant Biotechnol (Tokyo). 2022. PMID: 35800966 Free PMC article.
-
The Arabidopsis Rho of Plants GTPase ROP1 Is a Potential Calcium-Dependent Protein Kinase (CDPK) Substrate.Plants (Basel). 2021 Sep 29;10(10):2053. doi: 10.3390/plants10102053. Plants (Basel). 2021. PMID: 34685862 Free PMC article.
References
-
- Abo A, Pick E, Hall A, Totty N, Teahan CG, Segal AW (1991) Activation of the NADPH oxidase involves the small GTP-binding protein p21rac1. Nature 353: 668–670 - PubMed
-
- Akamatsu A, Wong HL, Fujiwara M, Okuda J, Nishide K, Uno K, Imai K, Umemura K, Kawasaki T, Kawano Y, et al. (2013) An OsCEBiP/OsCERK1-OsRacGEF1-OsRac1 module is an essential early component of chitin-induced rice immunity. Cell Host Microbe 13: 465–476 - PubMed
-
- Barbosa IC, Zourelidou M, Willige BC, Weller B, Schwechheimer C (2014) D6 PROTEIN KINASE activates auxin transport-dependent growth and PIN-FORMED phosphorylation at the plasma membrane. Dev Cell 29: 674–685 - PubMed
-
- Basu D, El-Assal Sel-D, Le J, Mallery EL, Szymanski DB (2004) Interchangeable functions of Arabidopsis PIROGI and the human WAVE complex subunit SRA1 during leaf epidermal development. Development 131: 4345–4355 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
