

Ml proteins from Mesorhizobium loti and MucR from Brucella abortus: an AT-rich core DNA-target site and oligomerization ability
- PMID: 29150637
- PMCID: PMC5693944
- DOI: 10.1038/s41598-017-16127-5
Ml proteins from Mesorhizobium loti and MucR from Brucella abortus: an AT-rich core DNA-target site and oligomerization ability
Abstract
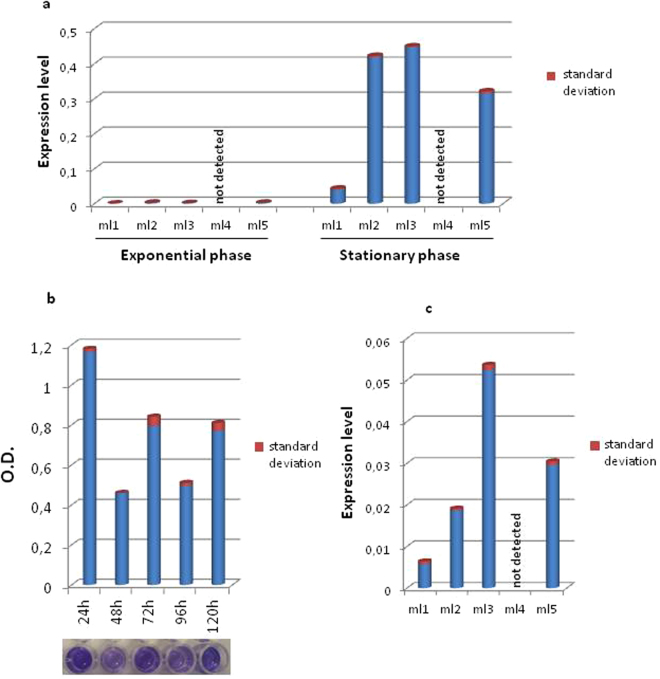
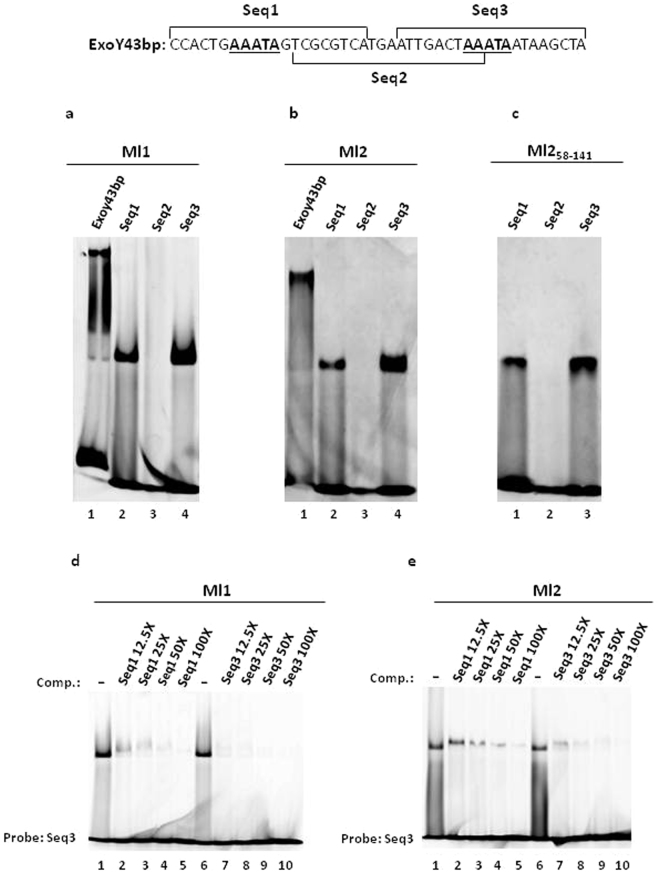
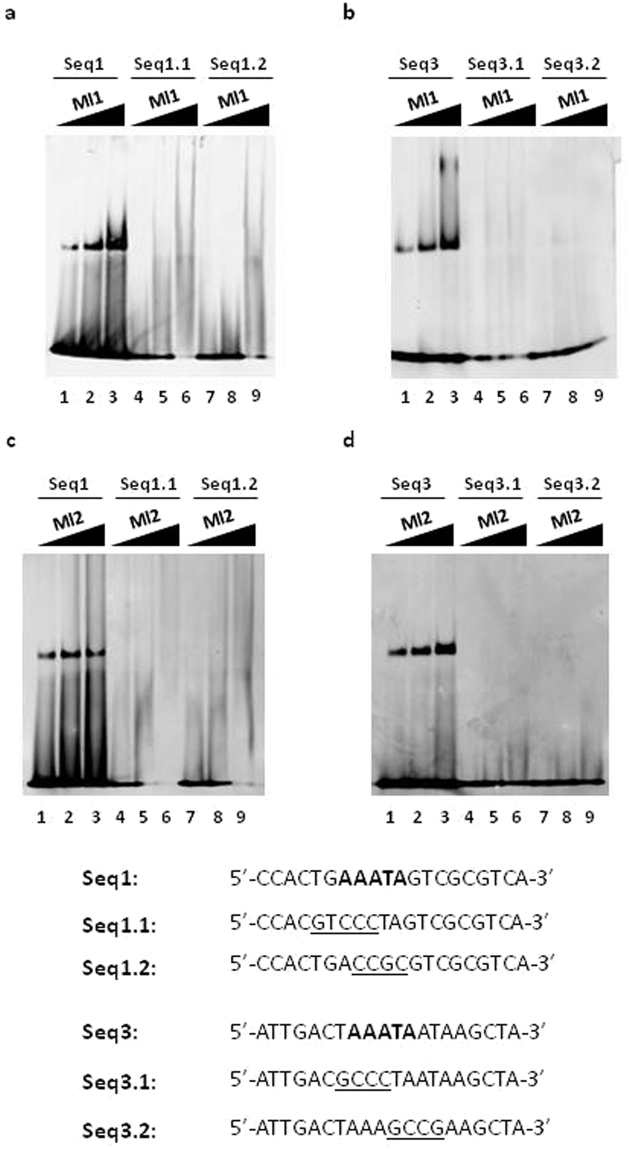
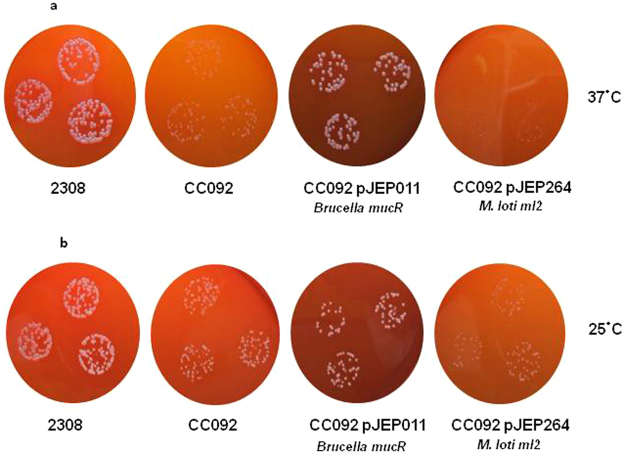
Mesorhizobium loti contains ten genes coding for proteins sharing high amino acid sequence identity with members of the Ros/MucR transcription factor family. Five of these Ros/MucR family members from Mesorhizobium loti (Ml proteins) have been recently structurally and functionally characterized demonstrating that Ml proteins are DNA-binding proteins. However, the DNA-binding studies were performed using the Ros DNA-binding site with the Ml proteins. Currently, there is no evidence as to when the Ml proteins are expressed during the Mesorhizobium lo ti life cycle as well as no information concerning their natural DNA-binding site. In this study, we examine the ml genes expression profile in Mesorhizobium loti and show that ml1, ml2, ml3 and ml5 are expressed during planktonic growth and in biofilms. DNA-binding experiments show that the Ml proteins studied bind a conserved AT-rich site in the promoter region of the exoY gene from Mesorhizobium loti and that the proteins make important contacts with the minor groove of DNA. Moreover, we demonstrate that the Ml proteins studied form higher-order oligomers through their N-terminal region and that the same AT-rich site is recognized by MucR from Brucella abortus using a similar mechanism involving contacts with the minor groove of DNA and oligomerization.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures



References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
