

Phosphatidylserine decarboxylase CT699, lysophospholipid acyltransferase CT775, and acyl-ACP synthase CT776 provide membrane lipid diversity to Chlamydia trachomatis
- PMID: 29150677
- PMCID: PMC5693948
- DOI: 10.1038/s41598-017-16116-8
Phosphatidylserine decarboxylase CT699, lysophospholipid acyltransferase CT775, and acyl-ACP synthase CT776 provide membrane lipid diversity to Chlamydia trachomatis
Abstract
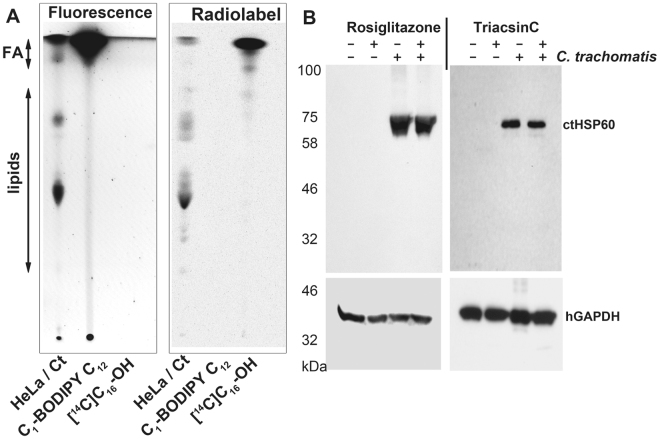
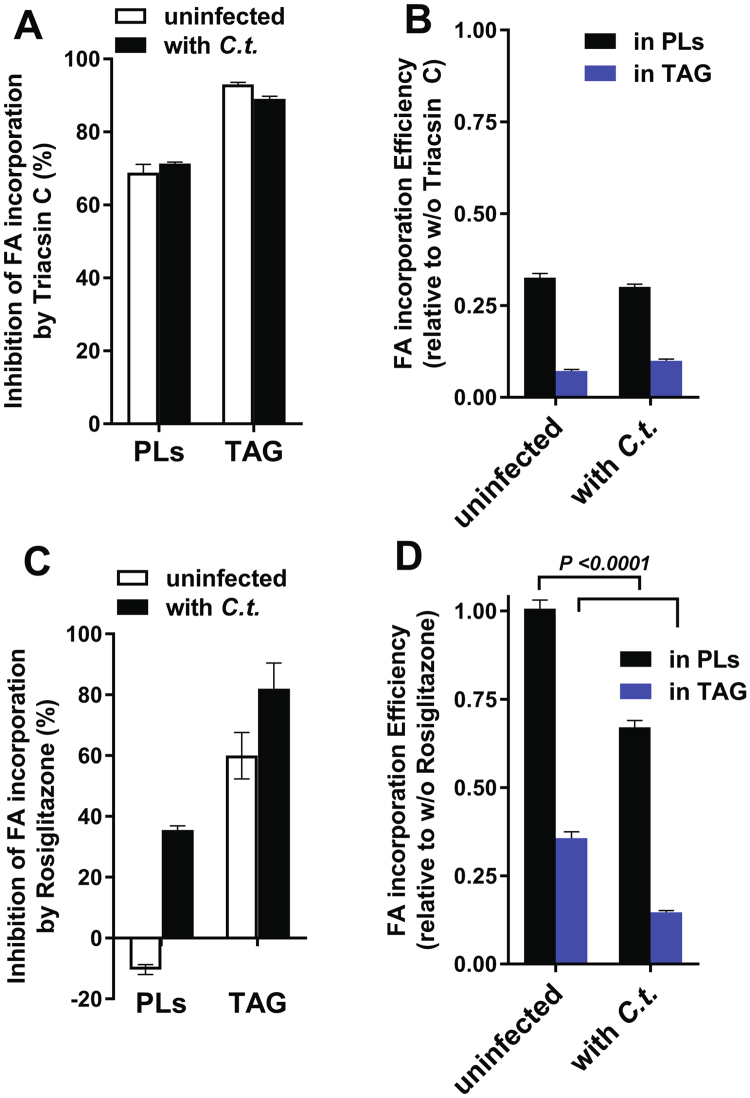
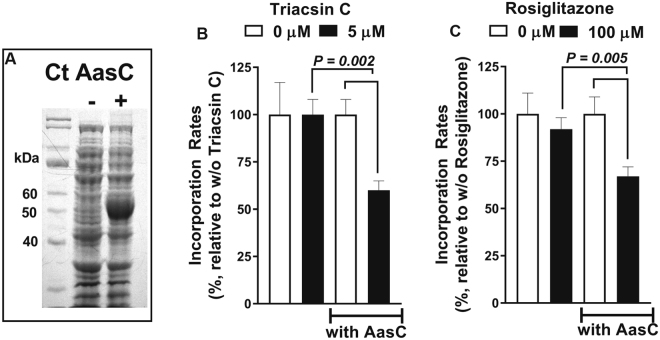
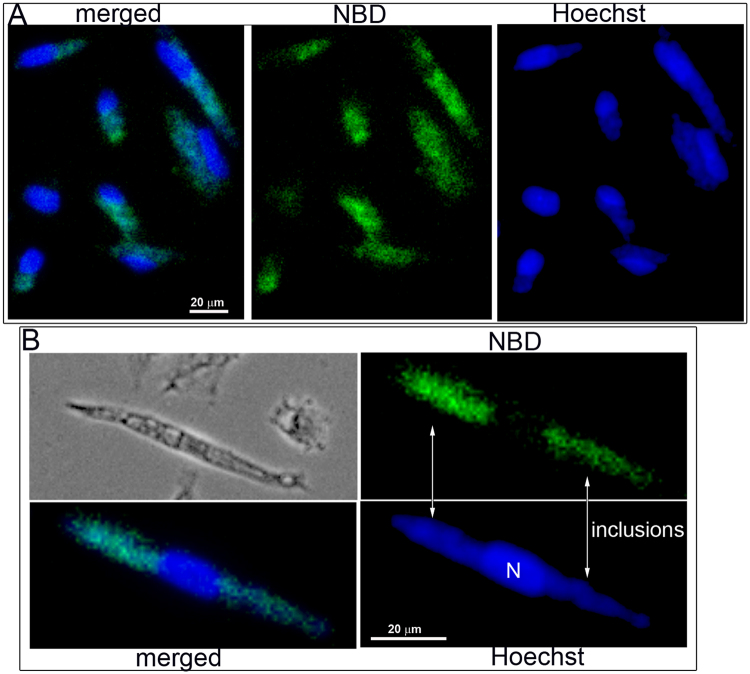
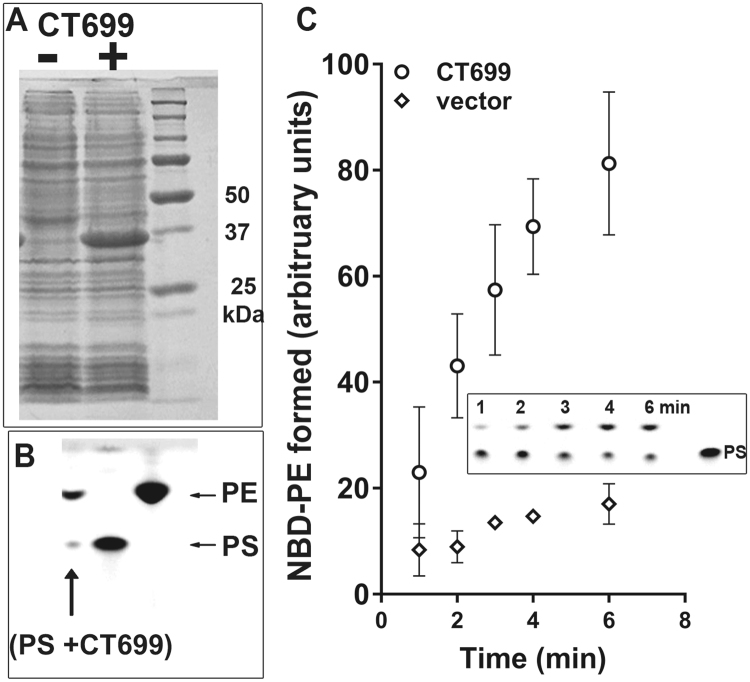
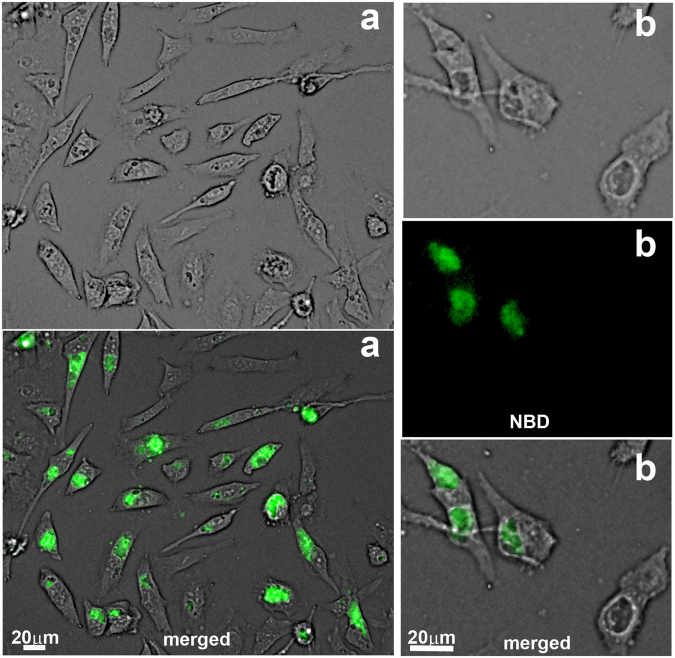
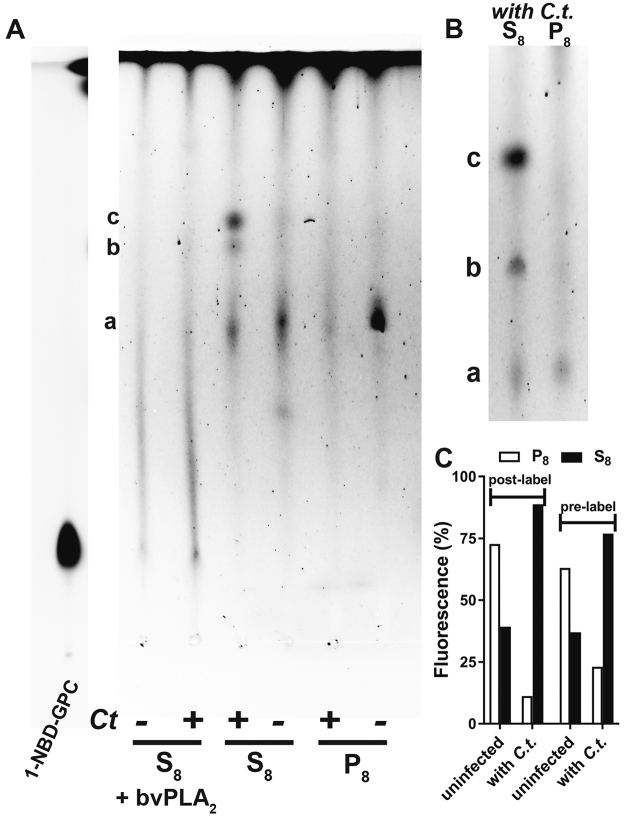
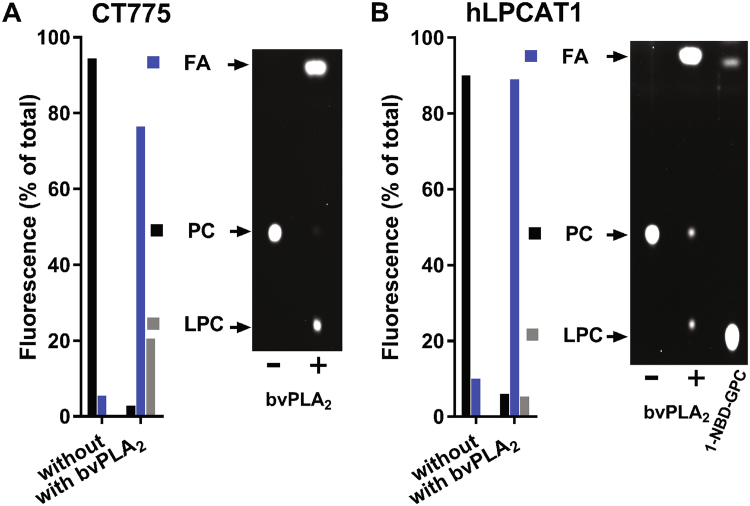
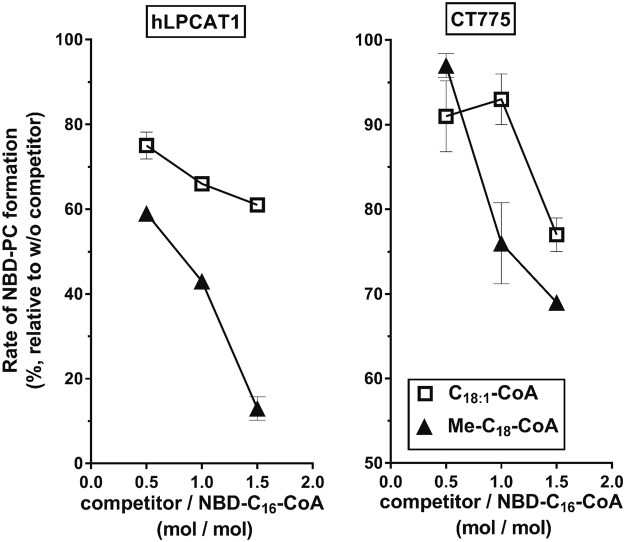
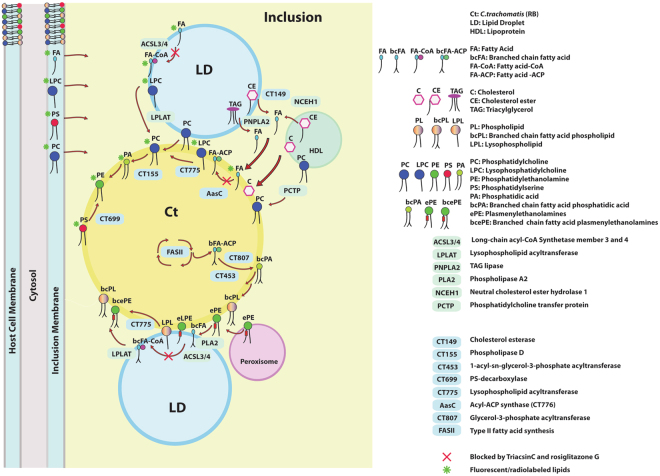
De novo lipid synthesis and scavenging of fatty acids (FA) are processes essential for the formation of the membrane of the human pathogen Chlamydia trachomatis (C.t.). Host FA are assimilated via esterification by the bacterial acyl-acyl carrier protein (ACP) synthase AasC but inhibitors of the host acyl-CoA synthetase enymes ACSL also impaired growth of C.t. in human cells. In E. coli, activity of AasC was sensitive to triacsin C and rosiglitazone G. The absence of a triacsin C-insensitive pathway and the increased inhibition by rosiglitazone G confirmed the sensitivity of the bacterial acyl-ACP synthase to these drugs in infected human cells. We found no evidence that the human ACSL enzymes are required for lipid formation by C.t. The broad substrate specificity of acyltransferase CT775 provides C.t. with the capacity to incorporate straight-chain and bacterial specific branched-chain fatty acids. CT775 accepts both acyl-ACP and acyl-CoA as acyl donors and, 1- or 2-acyl isomers of lysophosphoplipids as acyl acceptors. The enzyme responsible for remodeling of human phosphatidylserine to bacterial phosphatidylethanolamine was identified as CT699. These findings provide evidence that the pathogen has the ability to extend the lipid diversity of its membrane.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
