

When evolution is the solution to pollution: Key principles, and lessons from rapid repeated adaptation of killifish (Fundulus heteroclitus) populations
- PMID: 29151869
- PMCID: PMC5680427
- DOI: 10.1111/eva.12470
When evolution is the solution to pollution: Key principles, and lessons from rapid repeated adaptation of killifish (Fundulus heteroclitus) populations
Abstract
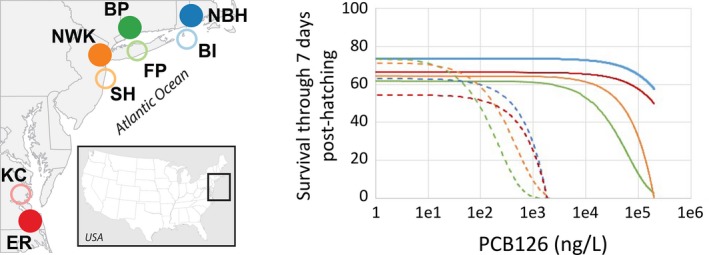
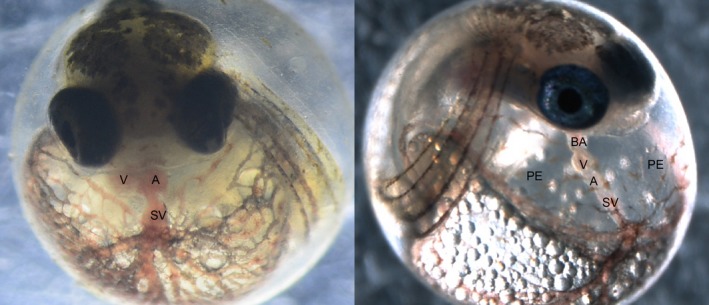
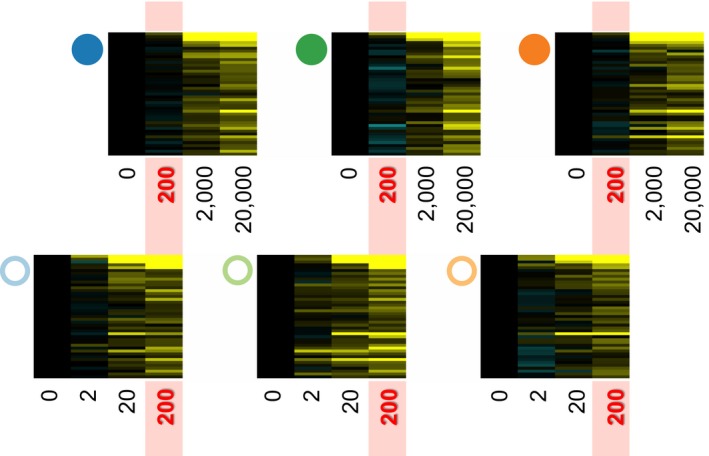
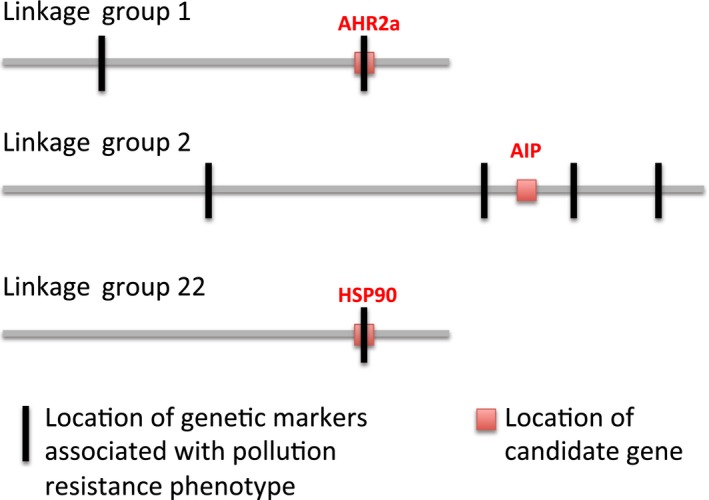
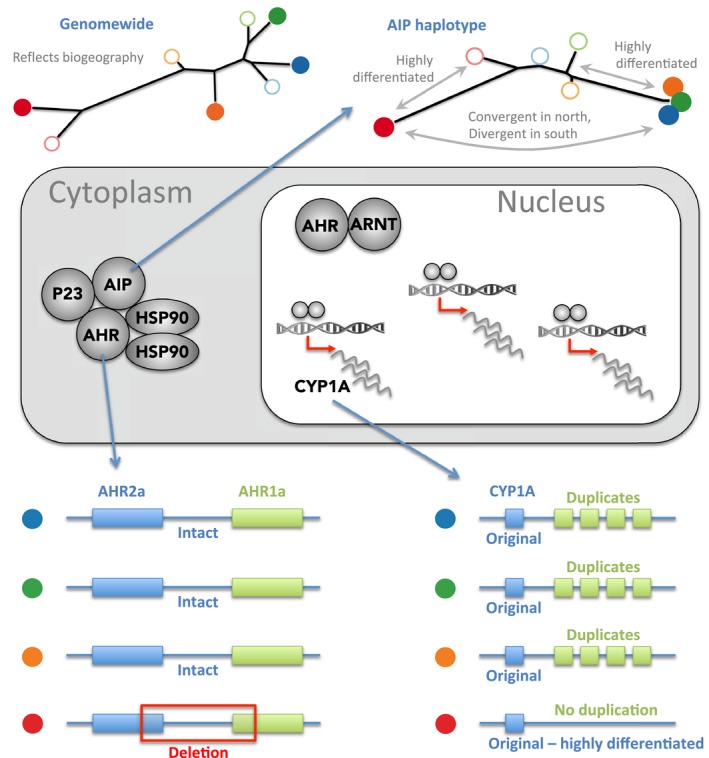
For most species, evolutionary adaptation is not expected to be sufficiently rapid to buffer the effects of human-mediated environmental changes, including environmental pollution. Here we review how key features of populations, the characteristics of environmental pollution, and the genetic architecture underlying adaptive traits, may interact to shape the likelihood of evolutionary rescue from pollution. Large populations of Atlantic killifish (Fundulus heteroclitus) persist in some of the most contaminated estuaries of the United States, and killifish studies have provided some of the first insights into the types of genomic changes that enable rapid evolutionary rescue from complexly degraded environments. We describe how selection by industrial pollutants and other stressors has acted on multiple populations of killifish and posit that extreme nucleotide diversity uniquely positions this species for successful evolutionary adaptation. Mechanistic studies have identified some of the genetic underpinnings of adaptation to a well-studied class of toxic pollutants; however, multiple genetic regions under selection in wild populations seem to reflect more complex responses to diverse native stressors and/or compensatory responses to primary adaptation. The discovery of these pollution-adapted killifish populations suggests that the evolutionary influence of anthropogenic stressors as selective agents occurs widely. Yet adaptation to chemical pollution in terrestrial and aquatic vertebrate wildlife may rarely be a successful "solution to pollution" because potentially adaptive phenotypes may be complex and incur fitness costs, and therefore be unlikely to evolve quickly enough, especially in species with small population sizes.
Keywords: adaptation; contemporary evolution; ecological genetics; ecotoxicology; genomics/proteomics; molecular evolution; natural selection and contemporary evolution; population genetics—empirical.
Figures
References
-
- Andersson, D. I. , & Hughes, D. (2010). Antibiotic resistance and its cost: Is it possible to reverse resistance? Nature Reviews Microbiology, 8, 260–271. - PubMed
-
- Andreev, D. , Kreitman, M. , Phillips, T. W. , Beeman, R. W. , & ffrench‐Constant, R. H. (1999). Multiple origins of cyclodiene insecticide resistance in Tribolium castaneum (Coleoptera: Tenebrionidae). Journal of Molecular Evolution, 48, 615–624. - PubMed
-
- Anselmo, H. M. R. , van den Berg, J. H. J. , Rietjens, I. M. C. M. , & Murk, A. J. (2012). Inhibition of cellular efflux pumps involved in multi xenobiotic resistance (MXR) in echinoid larvae as a possible mode of action for increased ecotoxicological risk of mixtures. Ecotoxicology, 21, 2276–2287. - PubMed
-
- Anthony, N. , Unruh, T. , Ganser, D. , & ffrench‐Constant, R. (1998). Duplication of the Rdl GABA receptor subunit gene in an insecticide‐resistant aphid, Myzus persicae . Molecular and General Genetics, 260, 165–175. - PubMed
-
- Antkiewicz, D. S. , Peterson, R. E. , & Heideman, W. (2006). Blocking expression of AHR2 and ARNT1 in zebrafish larvae protects against cardiac toxicity of 2,3,7,8‐tetrachlorodibenzo‐p‐dioxin. Toxicological Sciences, 94, 175–182. - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
