

Total Lipopolysaccharide from the Human Gut Microbiome Silences Toll-Like Receptor Signaling
- PMID: 29152585
- PMCID: PMC5686520
- DOI: 10.1128/mSystems.00046-17
Total Lipopolysaccharide from the Human Gut Microbiome Silences Toll-Like Receptor Signaling
Abstract
Cohabitation of microbial communities with the host enables the formation of a symbiotic relationship that maintains homeostasis in the gut and beyond. One prevailing model suggests that this relationship relies on the capacity of host cells and tissues to remain tolerant to the strong immune stimulation generated by the microbiota such as the activation of Toll-like receptor 4 (TLR4) pathways by lipopolysaccharide (LPS). Indeed, gut microbial LPS is thought to be one of the most potent activators of innate immune signaling and an important mediator of the microbiome's influence on host physiology. In this study, we performed computational and experimental analyses of healthy human fecal samples to examine the TLR4 signaling capacity of the gut microbiota. These analyses revealed that an immunoinhibitory activity of LPS, conserved across the members of the order Bacteroidales and derived from an underacylated structural feature, silences TLR4 signaling for the entire consortium of organisms inhabiting the human gut. Comparative analysis of metagenomic data from the Human Microbiome Project and healthy-donor samples indicates that immune silencing via LPS is a microbe-intrinsic feature in all healthy adults. These findings challenge the current belief that robust TLR4 signaling is a feature of the microbiome and demonstrate that microbiome-derived LPS has the ability to facilitate host tolerance of gut microbes. These findings have broad implications for how we model host-microbe interactions and for our understanding of microbiome-linked disease. IMPORTANCE While the ability for humans to host a complex microbial ecosystem is an essential property of life, the mechanisms allowing for immune tolerance of such a large microbial load are not completely understood and are currently the focus of intense research. This study shows that an important proinflammatory pathway that is commonly triggered by pathogenic bacteria upon interaction with the host is, in fact, actively repressed by the bacteria of the gut microbiome, supporting the idea that beneficial microbes themselves contribute to the immune tolerance in support of homeostasis. These findings are important for two reasons. First, many currently assume that proinflammatory signaling by lipopolysaccharide is a fundamental feature of the gut flora. This assumption influences greatly how host-microbiome interactions are theoretically modeled but also how they are experimentally studied, by using robust TLR signaling conditions to simulate commensals. Second, elucidation of the mechanisms that support host-microbe tolerance is key to the development of therapeutics for both intestinal and systemic inflammatory disorders.
Keywords: Toll-like receptors; lipopolysaccharide; microbial communities; microbiome; symbiosis; tolerance.
Figures
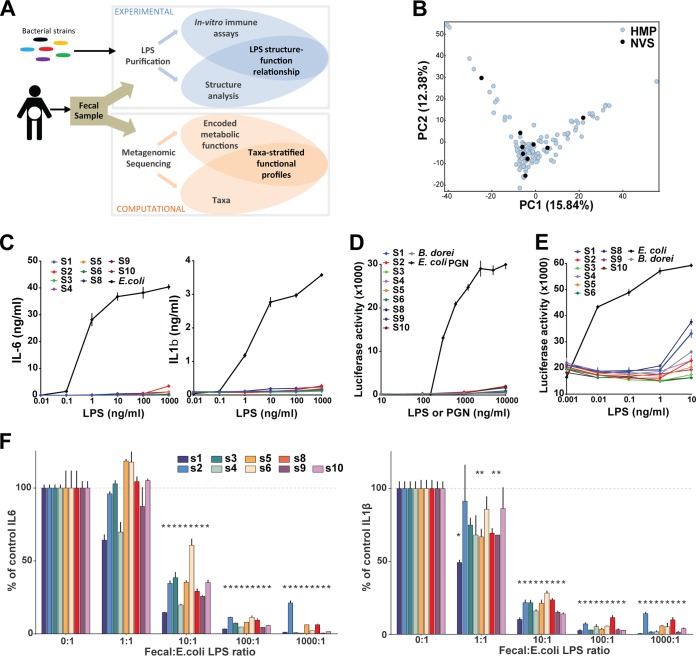
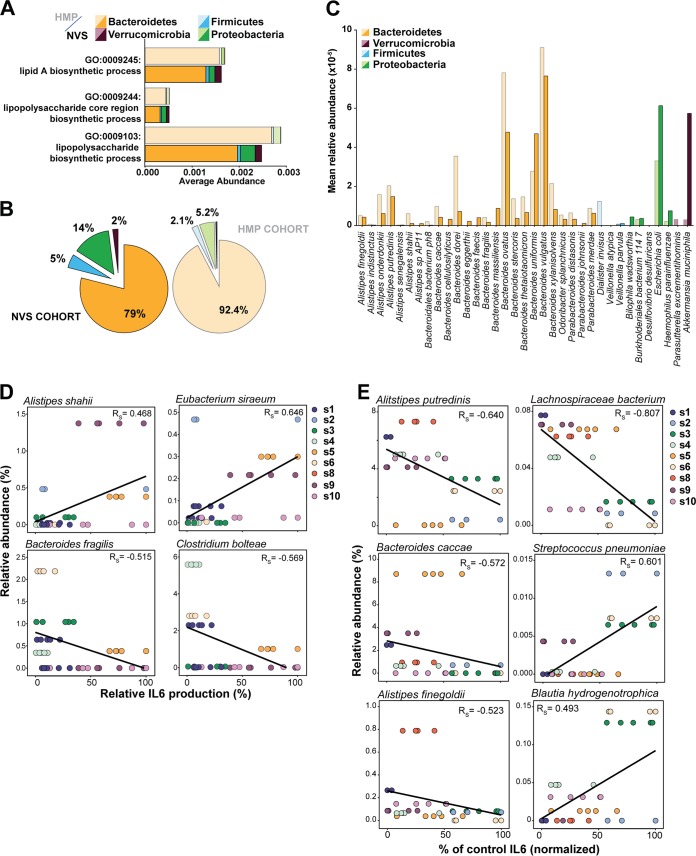
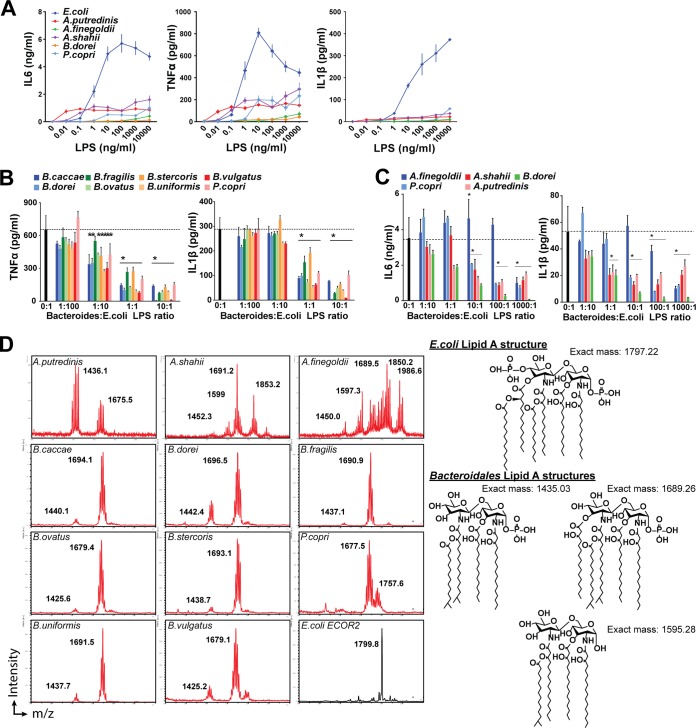
Comment in
-
Host response: Suppressing gut feelings.Nat Rev Microbiol. 2017 Dec 8;16(1):2. doi: 10.1038/nrmicro.2017.163. Nat Rev Microbiol. 2017. PMID: 29217843 No abstract available.
References
-
- Qin J, Li R, Raes J, Arumugam M, Burgdorf KS, Manichanh C, Nielsen T, Pons N, Levenez F, Yamada T, Mende DR, Li J, Xu J, Li S, Li D, Cao J, Wang B, Liang H, Zheng H, Xie Y, Tap J, Lepage P, Bertalan M, Batto JM, Hansen T, Le Paslier D, Linneberg A, Nielsen HB, Pelletier E, Renault P, Sicheritz-Ponten T, Turner K, Zhu H, Yu C, Li S, Jian M, Zhou Y, Li Y, Zhang X, Li S, Qin N, Yang H, Wang J, Brunak S, Dore J, Guarner F, Kristiansen K, Pedersen O, Parkhill J, Weissenbach J, Bork P, Ehrlich S, Wang J. 2010. A human gut microbial gene catalogue established by metagenomic sequencing. Nature 464:59–65. doi: 10.1038/nature08821. - DOI - PMC - PubMed
-
- Furusawa Y, Obata Y, Fukuda S, Endo TA, Nakato G, Takahashi D, Nakanishi Y, Uetake C, Kato K, Kato T, Takahashi M, Fukuda NN, Murakami S, Miyauchi E, Hino S, Atarashi K, Onawa S, Fujimura Y, Lockett T, Clarke JM, Topping DL, Tomita M, Hori S, Ohara O, Morita T, Koseki H, Kikuchi J, Honda K, Hase K, Ohno H. 2013. Commensal microbe-derived butyrate induces the differentiation of colonic regulatory T cells. Nature 504:446–450. doi: 10.1038/nature12721. - DOI - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
