

Polycomb Repressive Complex 2 Methylates Elongin A to Regulate Transcription
- PMID: 29153392
- PMCID: PMC5800316
- DOI: 10.1016/j.molcel.2017.10.025
Polycomb Repressive Complex 2 Methylates Elongin A to Regulate Transcription
Abstract
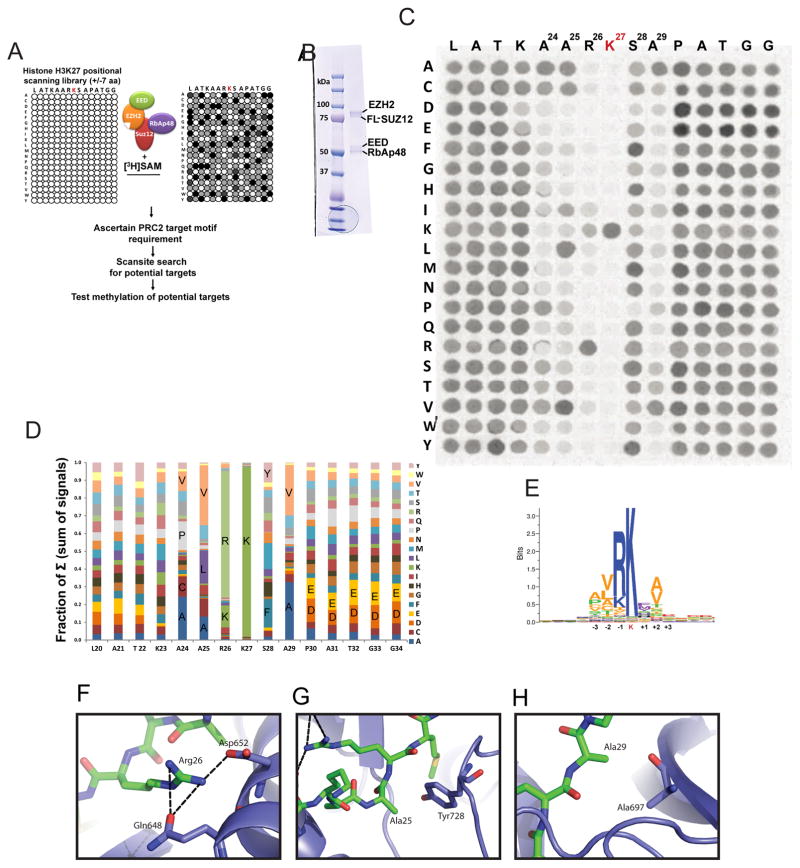
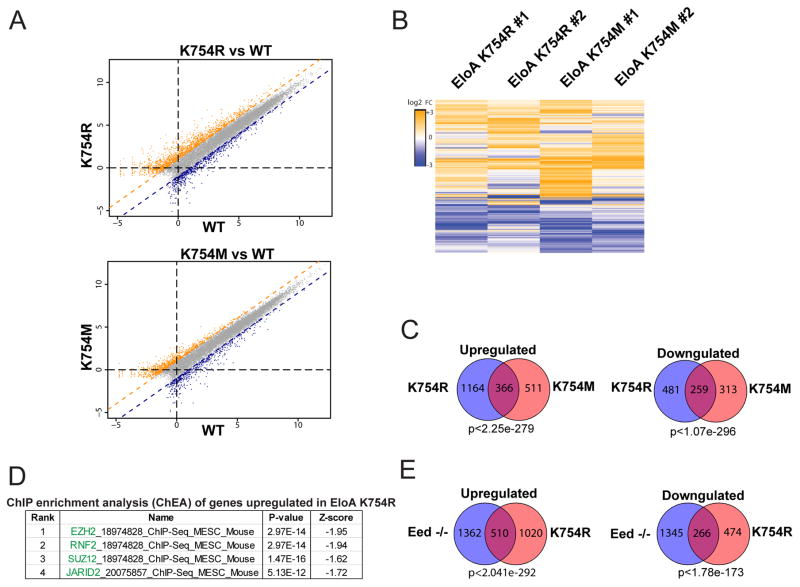
Polycomb repressive complex 2 (PRC2-EZH2) methylates histone H3 at lysine 27 (H3K27) and is required to maintain gene repression during development. Misregulation of PRC2 is linked to a range of neoplastic malignancies, which is believed to involve methylation of H3K27. However, the full spectrum of non-histone substrates of PRC2 that might also contribute to PRC2 function is not known. We characterized the target recognition specificity of the PRC2 active site and used the resultant data to screen for uncharacterized potential targets. The RNA polymerase II (Pol II) transcription elongation factor, Elongin A (EloA), is methylated by PRC2 in vivo. Mutation of the methylated EloA residue decreased repression of a subset of PRC2 target genes as measured by both steady-state and nascent RNA levels and perturbed embryonic stem cell differentiation. We propose that PRC2 modulates transcription of a subset of low expression target genes in part via methylation of EloA.
Keywords: EZH2; Elongin A; Elongin complex; Gene repression; Polycomb repressive complex 2 (PRC2); RNA polymerase II; SPOT assay; protein methylation; transcription.
Copyright © 2017 Elsevier Inc. All rights reserved.
Figures




References
-
- Abmayr SM, Carrozza MJ, Workman JL. Preparation of nuclear and cytoplasmic extracts from mammalian cells. Curr Protoc Pharmacol 2006 - PubMed
-
- Aso T, Lane WS, Conaway JW, Conaway RC. Elongin (SIII): a multisubunit regulator of elongation by RNA polymerase II. Science. 1995;269:1439–1443. - PubMed
-
- Beringer M, Pisano P, Di Carlo V, Blanco E, Chammas P, Vizán P, Gutiérrez A, Aranda S, Payer B, Wierer M, et al. EPOP Functionally Links Elongin and Polycomb in Pluripotent Stem Cells. Mol Cell. 2016;64:645–658. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
