

Vessel-Specific Reintroduction of CINNAMOYL-COA REDUCTASE1 (CCR1) in Dwarfed ccr1 Mutants Restores Vessel and Xylary Fiber Integrity and Increases Biomass
- PMID: 29158331
- PMCID: PMC5761799
- DOI: 10.1104/pp.17.01462
Vessel-Specific Reintroduction of CINNAMOYL-COA REDUCTASE1 (CCR1) in Dwarfed ccr1 Mutants Restores Vessel and Xylary Fiber Integrity and Increases Biomass
Abstract
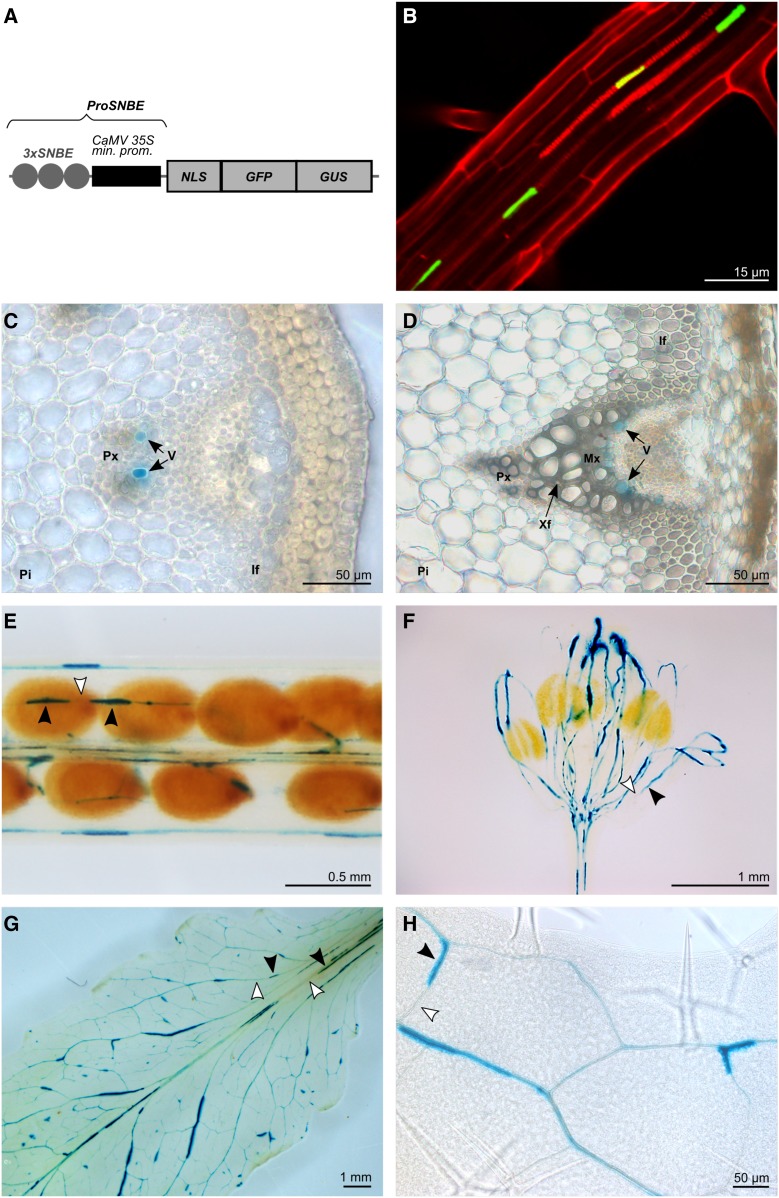
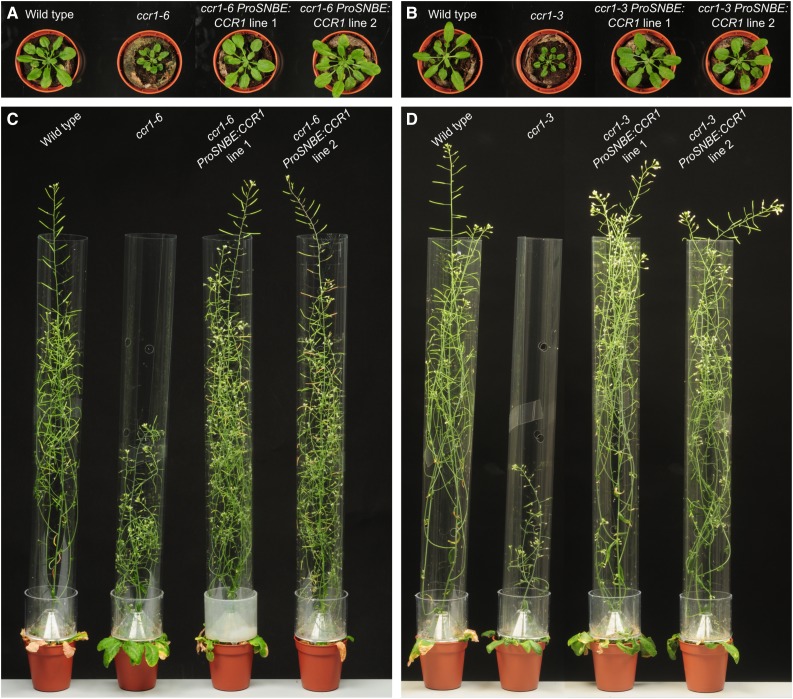
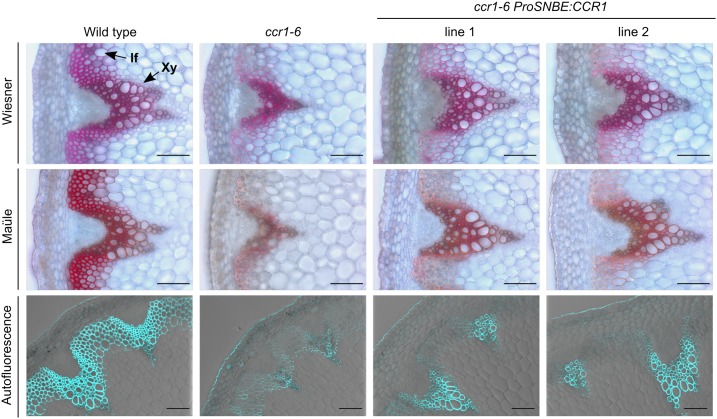
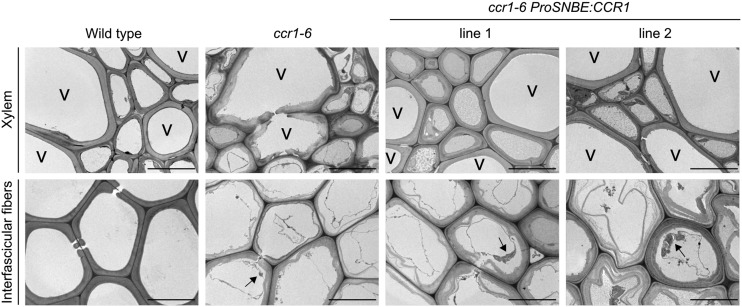
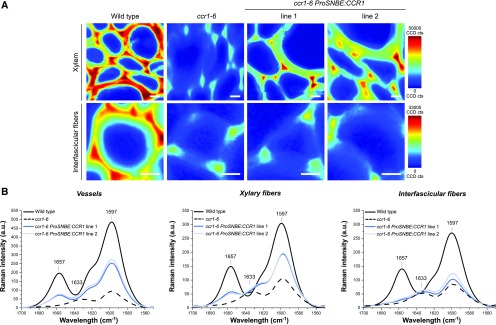
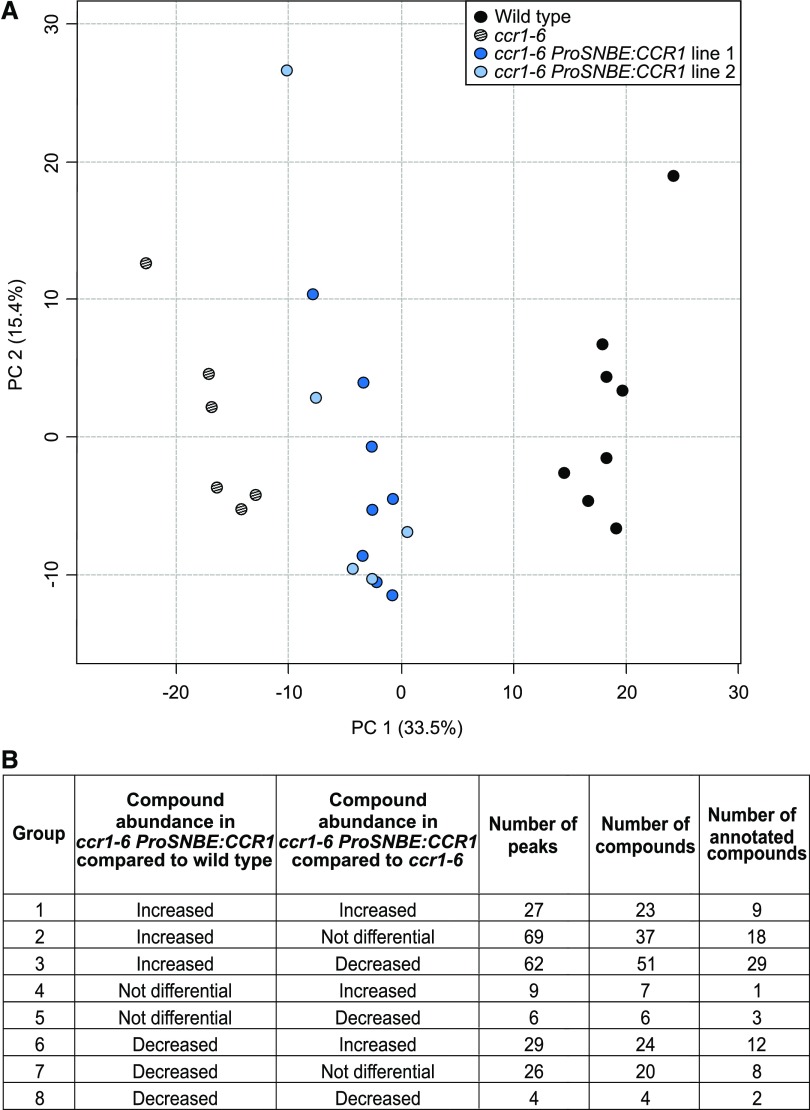
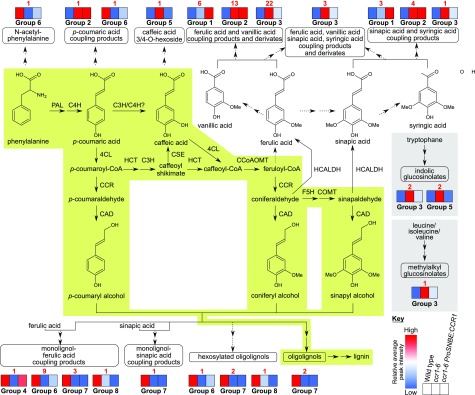
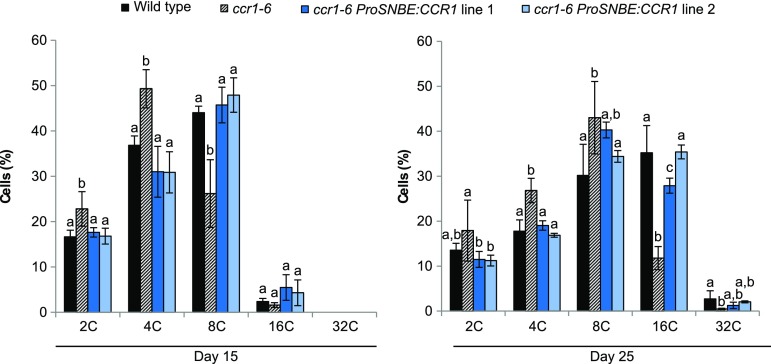
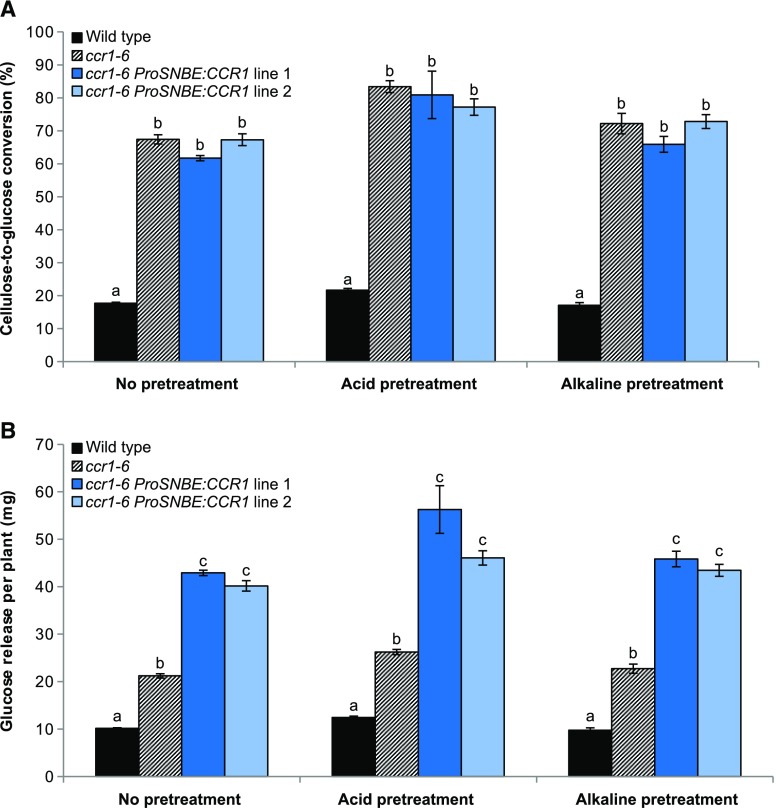
Lignocellulosic biomass is recalcitrant toward deconstruction into simple sugars due to the presence of lignin. To render lignocellulosic biomass a suitable feedstock for the bio-based economy, plants can be engineered to have decreased amounts of lignin. However, engineered plants with the lowest amounts of lignin exhibit collapsed vessels and yield penalties. Previous efforts were not able to fully overcome this phenotype without settling in sugar yield upon saccharification. Here, we reintroduced CINNAMOYL-COENZYME A REDUCTASE1 (CCR1) expression specifically in the protoxylem and metaxylem vessel cells of Arabidopsis (Arabidopsis thaliana) ccr1 mutants. The resulting ccr1 ProSNBE:CCR1 lines had overcome the vascular collapse and had a total stem biomass yield that was increased up to 59% as compared with the wild type. Raman analysis showed that monolignols synthesized in the vessels also contribute to the lignification of neighboring xylary fibers. The cell wall composition and metabolome of ccr1 ProSNBE:CCR1 still exhibited many similarities to those of ccr1 mutants, regardless of their yield increase. In contrast to a recent report, the yield penalty of ccr1 mutants was not caused by ferulic acid accumulation but was (largely) the consequence of collapsed vessels. Finally, ccr1 ProSNBE:CCR1 plants had a 4-fold increase in total sugar yield when compared with wild-type plants.
© 2018 American Society of Plant Biologists. All Rights Reserved.
Figures

References
-
- Agarwal UP, Atalla RH (1990) Formation and identification of cis/trans ferulic acid in photoyellowed white spruce mechanical pulp. J Wood Chem Technol 10: 169–190
-
- Agarwal UP, McSweeny JD, Ralph SA (2011) FT-Raman investigation of milled-wood lignins: softwood, hardwood, and chemically modified black spruce lignins. J Wood Chem Technol 31: 324–344
-
- Agarwal UP, Ralph SA, Atalla RH (1997) FT Raman spectroscopic study of softwood lignin. In 9th International Symposium on Wood and Pulping Chemistry: Proceedings, Montréal, Québec, 1997.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
