

Ethylene Receptors Signal via a Noncanonical Pathway to Regulate Abscisic Acid Responses
- PMID: 29158332
- PMCID: PMC5761792
- DOI: 10.1104/pp.17.01321
Ethylene Receptors Signal via a Noncanonical Pathway to Regulate Abscisic Acid Responses
Abstract
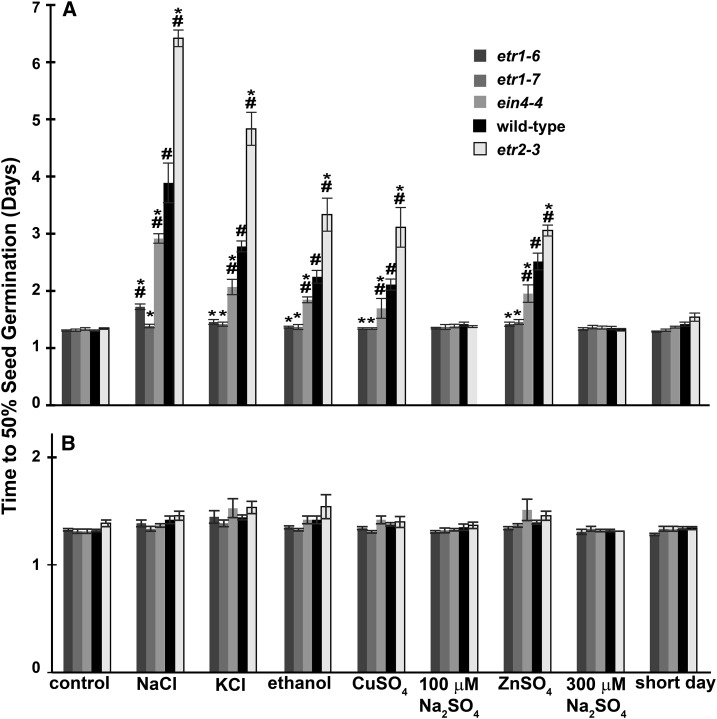
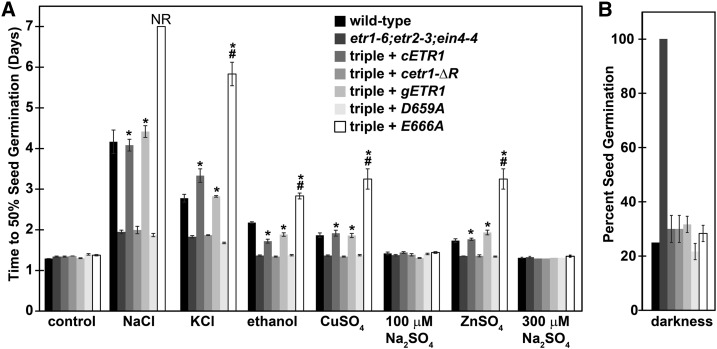
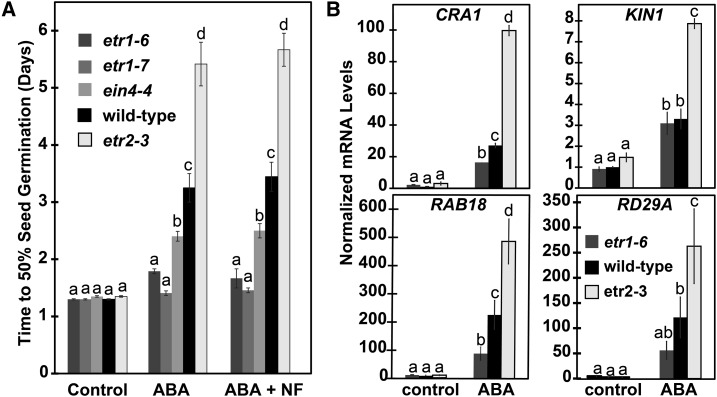
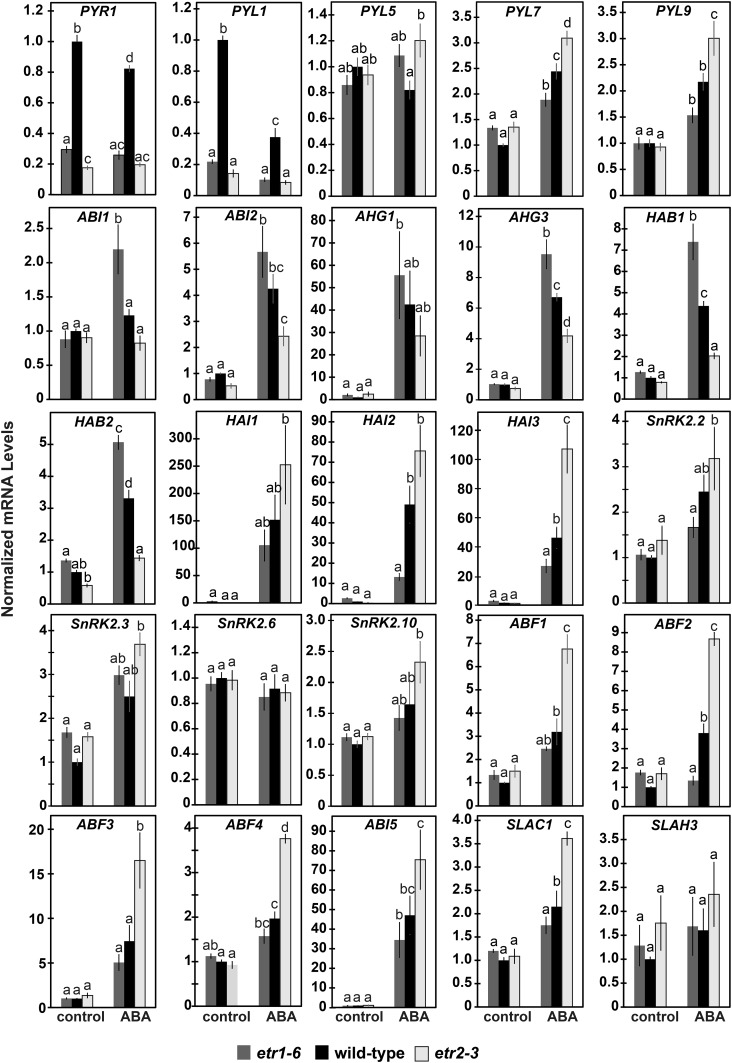
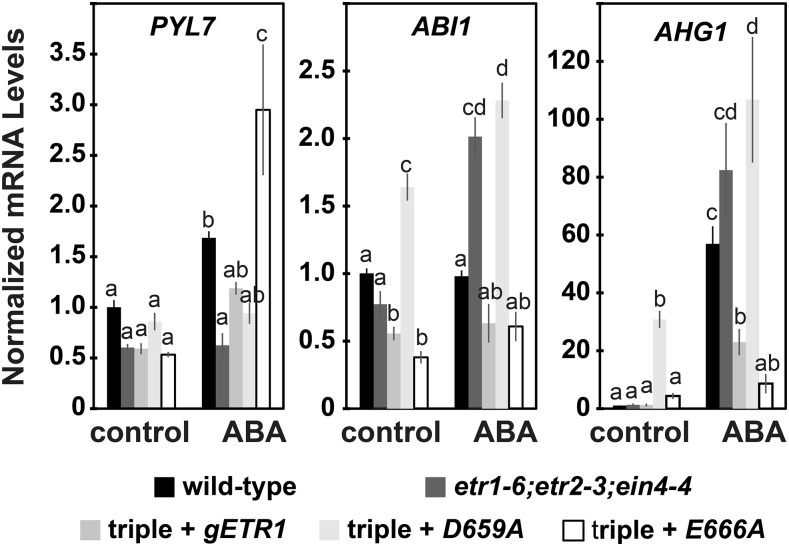
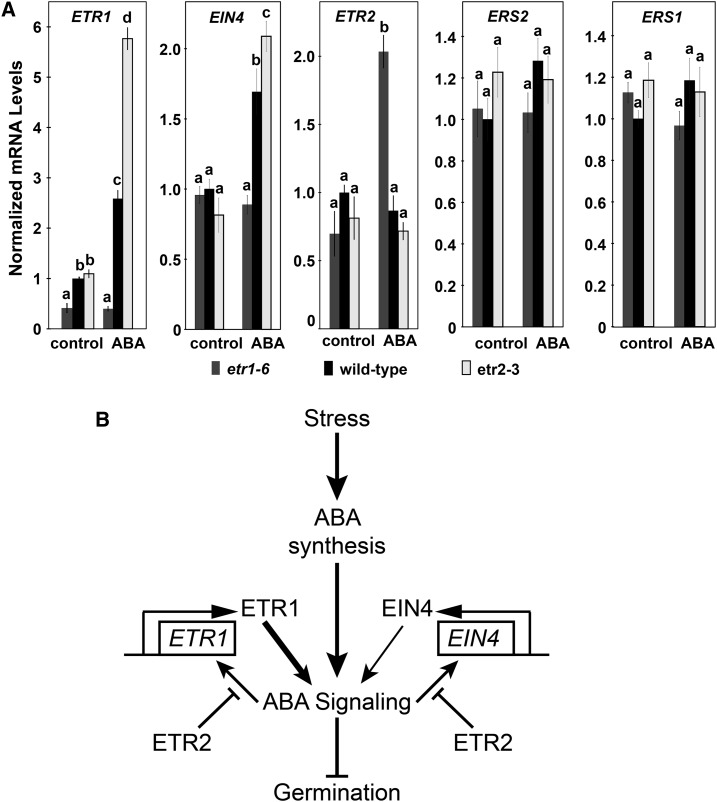
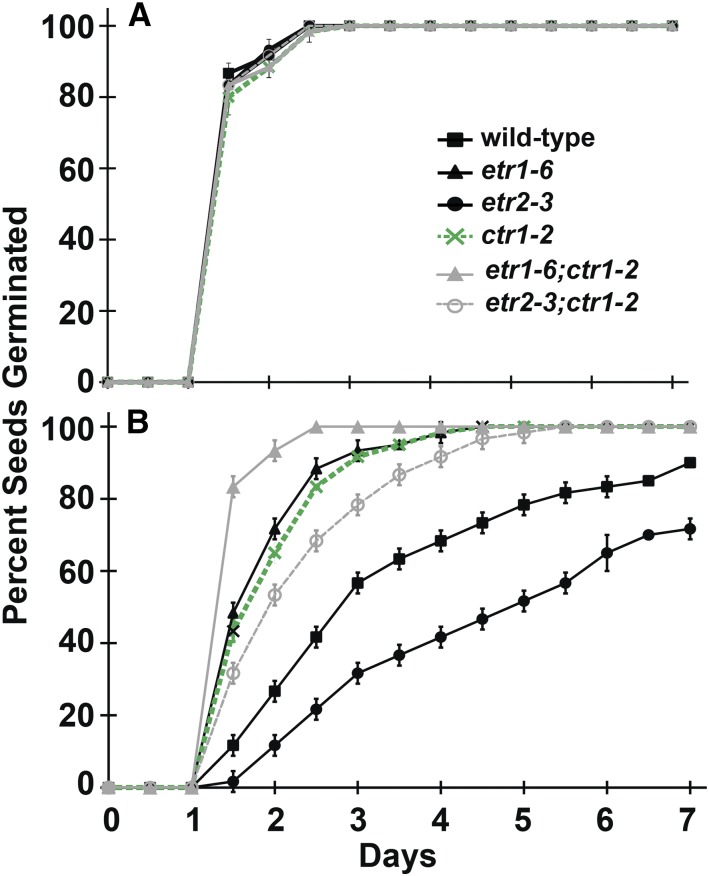
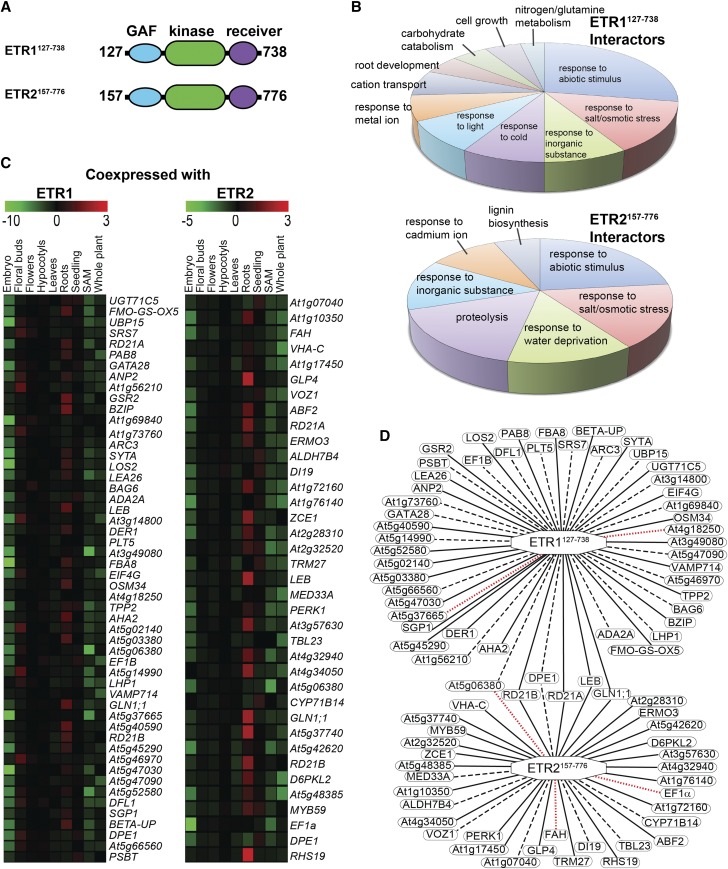
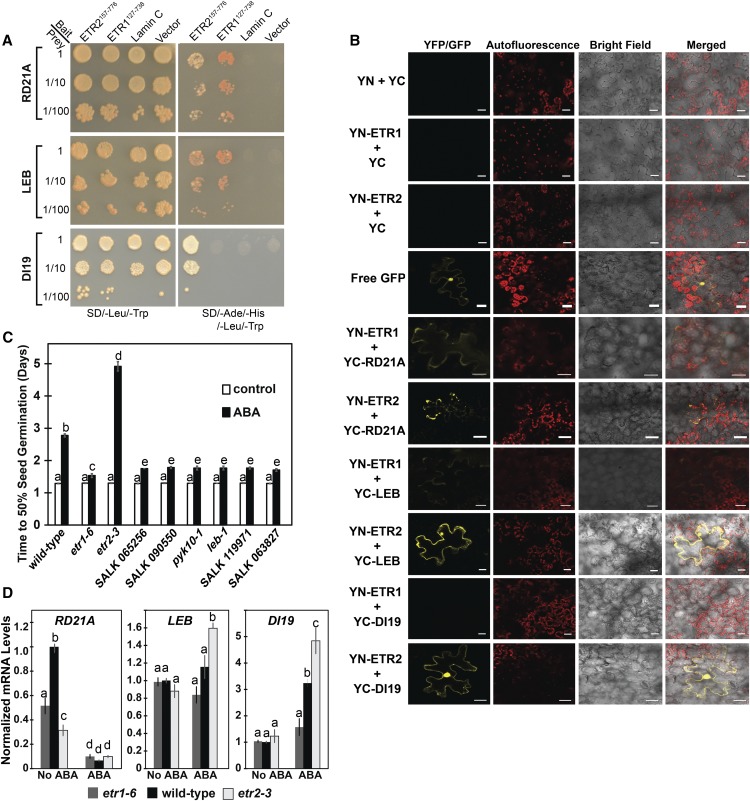
Ethylene is a gaseous plant hormone perceived by a family of receptors in Arabidopsis (Arabidopsis thaliana) including ETHYLENE RESPONSE1 (ETR1) and ETR2. Previously we showed that etr1-6 loss-of-function plants germinate better and etr2-3 loss-of-function plants germinate worse than wild-type under NaCl stress and in response to abscisic acid (ABA). In this study, we expanded these results by showing that ETR1 and ETR2 have contrasting roles in the control of germination under a variety of inhibitory conditions for seed germination such as treatment with KCl, CuSO4, ZnSO4, and ethanol. Pharmacological and molecular biology results support a model where ETR1 and ETR2 are indirectly affecting the expression of genes encoding ABA signaling proteins to affect ABA sensitivity. The receiver domain of ETR1 is involved in this function in germination under these conditions and controlling the expression of genes encoding ABA signaling proteins. Epistasis analysis demonstrated that these contrasting roles of ETR1 and ETR2 do not require the canonical ethylene signaling pathway. To explore the importance of receptor-protein interactions, we conducted yeast two-hybrid screens using the cytosolic domains of ETR1 and ETR2 as bait. Unique interacting partners with either ETR1 or ETR2 were identified. We focused on three of these proteins and confirmed the interactions with receptors. Loss of these proteins led to faster germination in response to ABA, showing that they are involved in ABA responses. Thus, ETR1 and ETR2 have both ethylene-dependent and -independent roles in plant cells that affect responses to ABA.
© 2018 American Society of Plant Biologists. All Rights Reserved.
Figures
References
-
- Abeles F, Morgan P, Saltveit MJ (1992) Ethylene in Plant Biology, 2nd Ed. Academic Press, San Diego, CA
-
- Alonso JM, Hirayama T, Roman G, Nourizadeh S, Ecker JR (1999) EIN2, a bifunctional transducer of ethylene and stress responses in Arabidopsis. Science 284: 2148–2152 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
