

MiR-29b antagonizes the pro-inflammatory tumor-promoting activity of multiple myeloma-educated dendritic cells
- PMID: 29158557
- PMCID: PMC5886056
- DOI: 10.1038/leu.2017.336
MiR-29b antagonizes the pro-inflammatory tumor-promoting activity of multiple myeloma-educated dendritic cells
Abstract
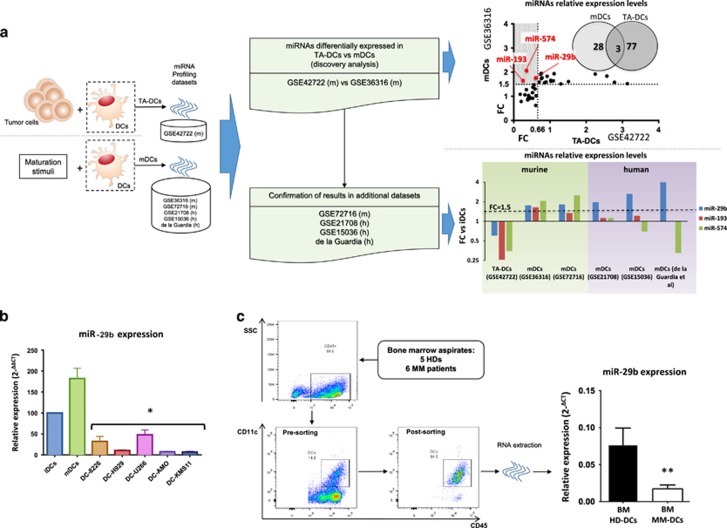
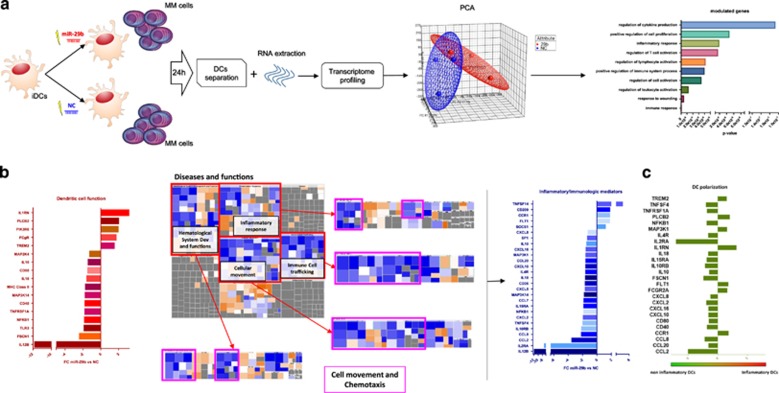
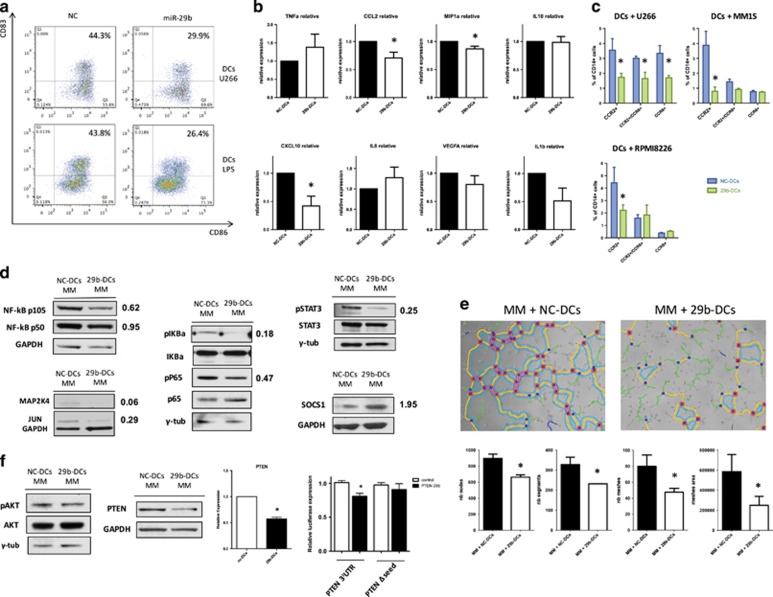
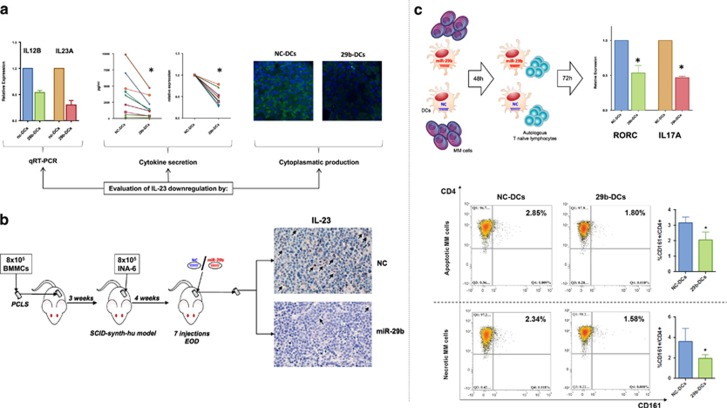
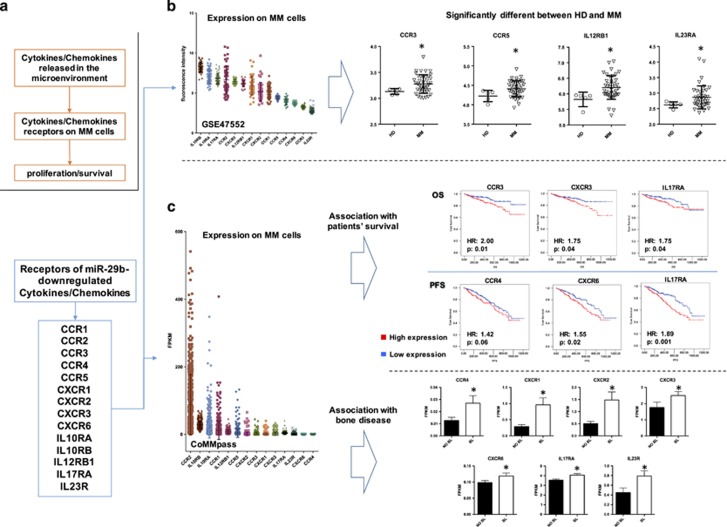
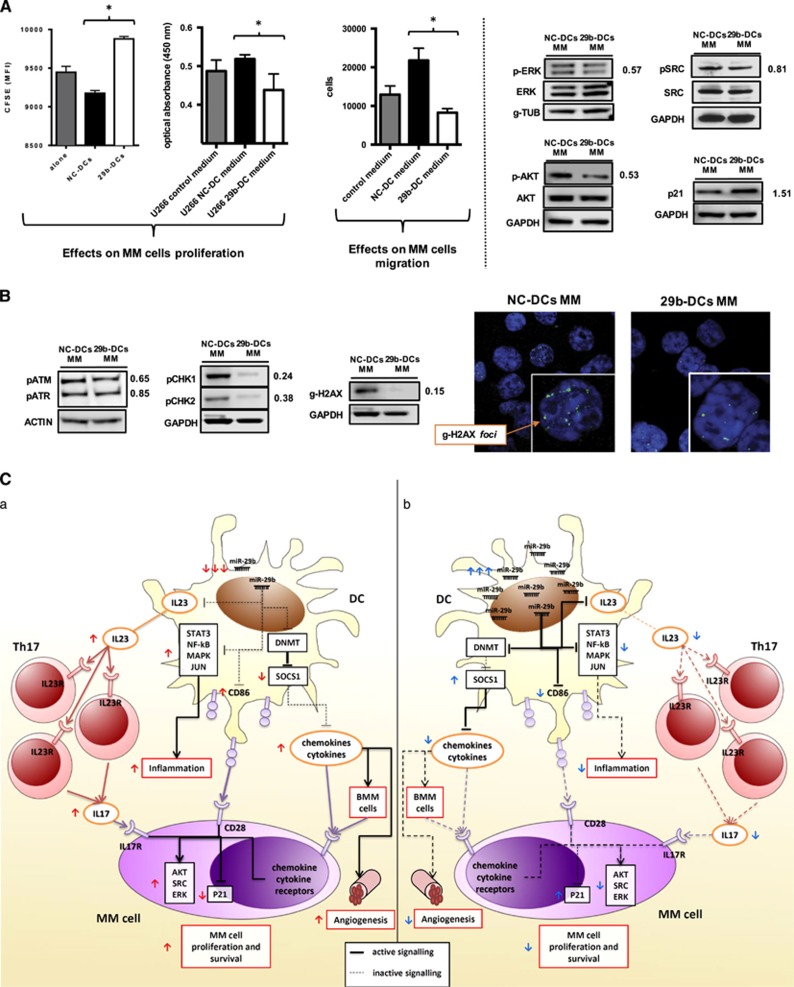
Dendritic cells (DCs) have a key role in regulating tumor immunity, tumor cell growth and drug resistance. We hypothesized that multiple myeloma (MM) cells might recruit and reprogram DCs to a tumor-permissive phenotype by changes within their microRNA (miRNA) network. By analyzing six different miRNA-profiling data sets, miR-29b was identified as the only miRNA upregulated in normal mature DCs and significantly downregulated in tumor-associated DCs. This finding was validated in primary DCs co-cultured in vitro with MM cell lines and in primary bone marrow DCs from MM patients. In DCs co-cultured with MM cells, enforced expression of miR-29b counteracted pro-inflammatory pathways, including signal transducer and activator of transcription 3 and nuclear factor-κB, and cytokine/chemokine signaling networks, which correlated with patients' adverse prognosis and development of bone disease. Moreover, miR-29b downregulated interleukin-23 in vitro and in the SCID-synth-hu in vivo model, and antagonized a Th17 inflammatory response. All together, these effects translated into strong anti-proliferative activity and reduction of genomic instability of MM cells. Our study demonstrates that MM reprograms the DCs functional phenotype by downregulating miR-29b whose reconstitution impairs DCs ability to sustain MM cell growth and survival. These results underscore miR-29b as an innovative and attractive candidate for miRNA-based immune therapy of MM.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
miR-29b defines the pro-/anti-proliferative effects of S100A7 in breast cancer.Mol Cancer. 2015 Jan 27;14:11. doi: 10.1186/s12943-014-0275-z. Mol Cancer. 2015. PMID: 25622979 Free PMC article.
-
Overexpression of microRNA-29b induces apoptosis of multiple myeloma cells through down regulating Mcl-1.Biochem Biophys Res Commun. 2011 Oct 14;414(1):233-9. doi: 10.1016/j.bbrc.2011.09.063. Epub 2011 Sep 17. Biochem Biophys Res Commun. 2011. PMID: 21951844
-
Genistein inhibits the proliferation of human multiple myeloma cells through suppression of nuclear factor-κB and upregulation of microRNA-29b.Mol Med Rep. 2016 Feb;13(2):1627-32. doi: 10.3892/mmr.2015.4740. Epub 2015 Dec 30. Mol Med Rep. 2016. PMID: 26718793
-
Regulation of the MIR155 host gene in physiological and pathological processes.Gene. 2013 Dec 10;532(1):1-12. doi: 10.1016/j.gene.2012.12.009. Epub 2012 Dec 14. Gene. 2013. PMID: 23246696 Review.
-
Promises and challenges of MicroRNA-based treatment of multiple myeloma.Curr Cancer Drug Targets. 2012 Sep;12(7):838-46. doi: 10.2174/156800912802429355. Curr Cancer Drug Targets. 2012. PMID: 22671926 Free PMC article. Review.
Cited by
-
Multiple Myeloma Bone Disease: Implication of MicroRNAs in Its Molecular Background.Int J Mol Sci. 2021 Feb 27;22(5):2375. doi: 10.3390/ijms22052375. Int J Mol Sci. 2021. PMID: 33673480 Free PMC article. Review.
-
The Role and Function of microRNA in the Pathogenesis of Multiple Myeloma.Cancers (Basel). 2019 Nov 6;11(11):1738. doi: 10.3390/cancers11111738. Cancers (Basel). 2019. PMID: 31698726 Free PMC article. Review.
-
Mechanisms of Immune Evasion in Multiple Myeloma: Open Questions and Therapeutic Opportunities.Cancers (Basel). 2021 Jun 28;13(13):3213. doi: 10.3390/cancers13133213. Cancers (Basel). 2021. PMID: 34203150 Free PMC article. Review.
-
MicroRNA in cancer therapy: breakthroughs and challenges in early clinical applications.J Exp Clin Cancer Res. 2025 Apr 21;44(1):126. doi: 10.1186/s13046-025-03391-x. J Exp Clin Cancer Res. 2025. PMID: 40259326 Free PMC article. Review.
-
Multiple myeloma: the (r)evolution of current therapy and a glance into future.Haematologica. 2020 Oct 1;105(10):2358-2367. doi: 10.3324/haematol.2020.247015. Haematologica. 2020. PMID: 33054076 Free PMC article.
References
-
- Rossi M, Botta C, Correale P, Tassone P, Tagliaferri P. Immunologic microenvironment and personalized treatment in multiple myeloma. Expert Opin Biol Ther 2013; 13 (Suppl 1): S83–93. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
