

Insertion of the Type-I IFN Decoy Receptor B18R in a miRNA-Tagged Semliki Forest Virus Improves Oncolytic Capacity but Results in Neurotoxicity
- PMID: 29159280
- PMCID: PMC5684435
- DOI: 10.1016/j.omto.2017.10.001
Insertion of the Type-I IFN Decoy Receptor B18R in a miRNA-Tagged Semliki Forest Virus Improves Oncolytic Capacity but Results in Neurotoxicity
Abstract
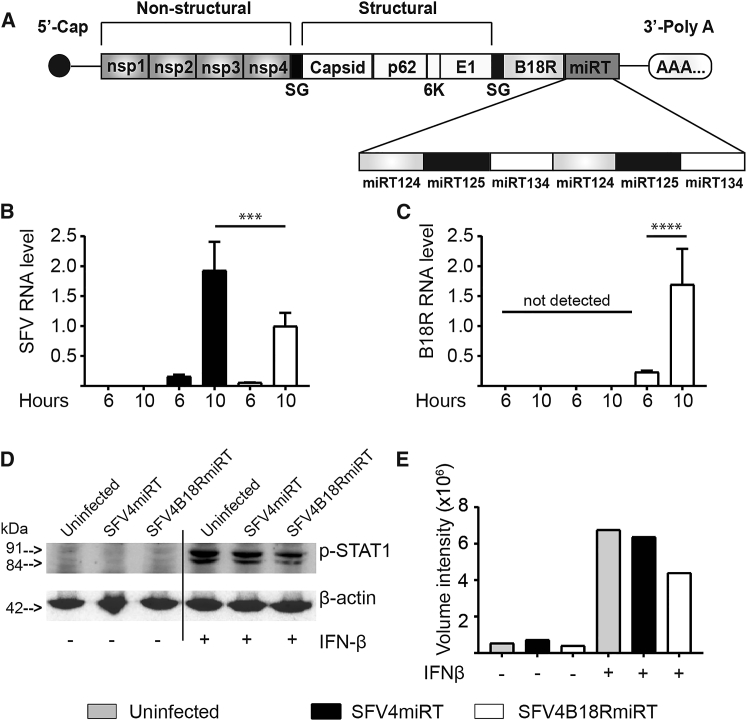
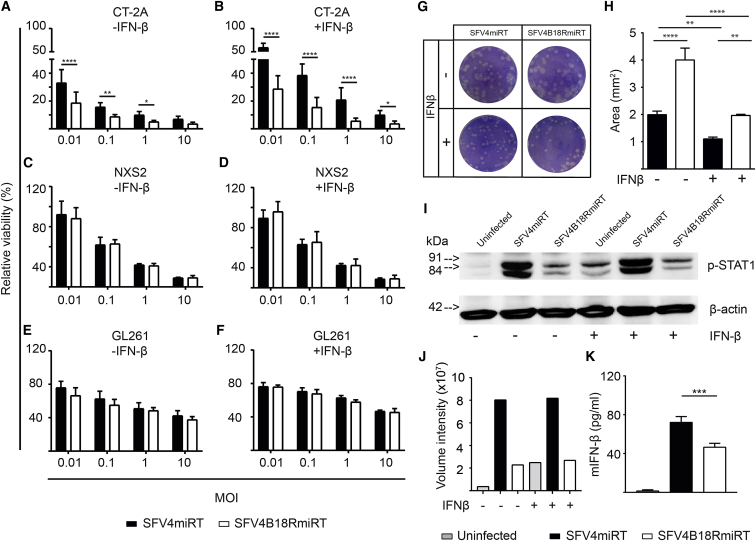
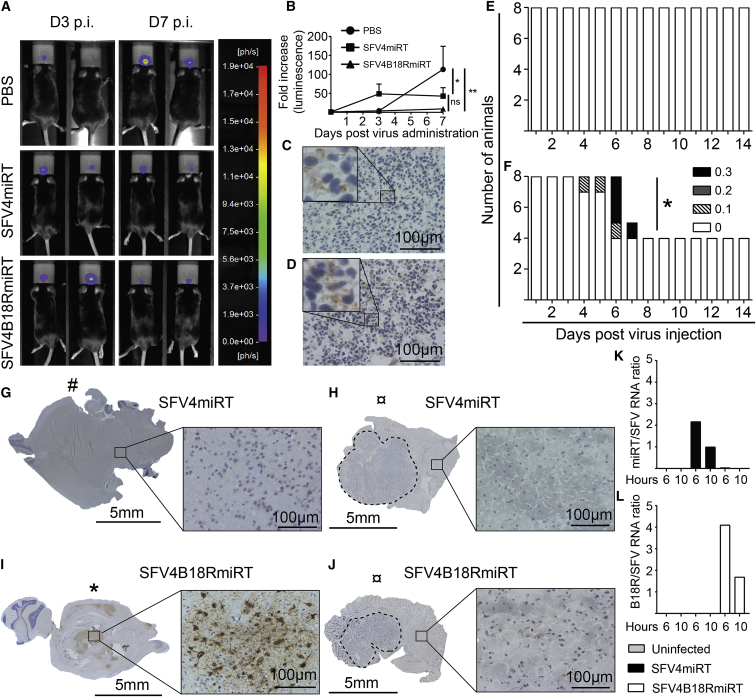
Oncolytic Semliki Forest virus (SFV) has been suggested as a potential candidate for the treatment of glioblastoma and neuroblastoma. However, the oncolytic capacity of SFV is restricted by the anti-viral type-I interferon (IFN) response. The aim of this study was to increase the oncolytic capacity of a microRNA target tagged SFV against glioblastoma by arming it with the Vaccinia-virus-encoded type-I IFN decoy receptor B18R (SFV4B18RmiRT) to neutralize type-I IFN response. Expression of B18R by SFV4B18RmiRT aided neutralization of IFN-β, which was shown by reduced STAT-1 phosphorylation and improved virus spread in plaque assays. B18R expression by SFV4 increased its oncolytic capacity in vitro against murine glioblastoma (CT-2A), regardless of the presence of exogenous IFN-β. Both SFV4B18RmiRT and SFV4miRT treatments controlled tumor growth in mice with syngeneic orthotopic gliomablastoma (CT-2A). However, treatment with SFV4B18RmiRT induced severe neurological symptoms in some mice because of virus replication in the healthy brain. Neither neurotoxicity nor virus replication in the brain was observed when SFV4miRT was administered. In summary, our results indicate that the oncolytic capacity of SFV4 was improved in vitro and in vivo by incorporation of B18R, but neurotoxicity of the virus was increased, possibly due to loss of microRNA targets.
Keywords: B18R; SFV; Semliki Forest virus; glioblastoma; type I interferon.
Figures
References
-
- Alifieris C., Trafalis D.T. Glioblastoma multiforme: pathogenesis and treatment. Pharmacol. Ther. 2015;152:63–82. - PubMed
-
- Furnari F.B., Fenton T., Bachoo R.M., Mukasa A., Stommel J.M., Stegh A., Hahn W.C., Ligon K.L., Louis D.N., Brennan C. Malignant astrocytic glioma: genetics, biology, and paths to treatment. Genes Dev. 2007;21:2683–2710. - PubMed
-
- Fazakerley J.K. Pathogenesis of Semliki Forest virus encephalitis. J. Neurovirol. 2002;8(Suppl 2):66–74. - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
